Rhodopsin
From Proteopedia
(Difference between revisions)
m (fixed so no table of contents show now that list out of structure section) |
|||
| (60 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
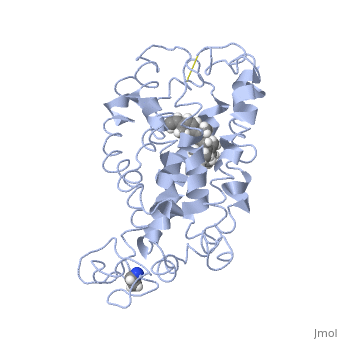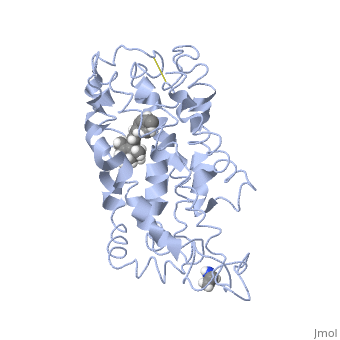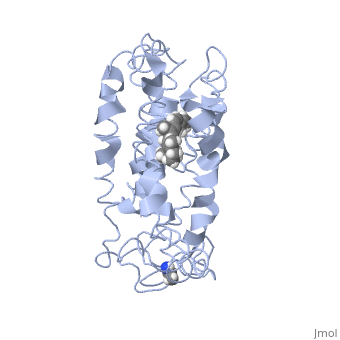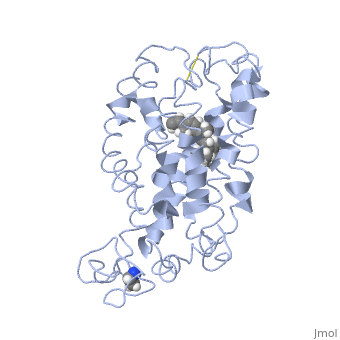
| - | <StructureSection load=' | + | <StructureSection load='1jfp' size='350' side='right' scene='' caption='Bovine rhodopsin complex with retinal (PDB code [[1jfp]])'> |
| + | |||
==Introduction== | ==Introduction== | ||
===Rhodopsin=== | ===Rhodopsin=== | ||
| - | Rhodopsin, a homodimeric protein, is a highly characterized [[G protein-coupled receptor]] found in membranous disks of the outer segments of rod and cone cells, though rhodopsin is more concentrated in rod cells which are sensitive to light but cannot discriminate colors. Rhodopsin is part of the superfamily of G protein-coupled receptors that mediate responses to visual, olfactory, hormonal, and neurotransmitter signals among others<ref name="Article1">PMID:20004206</ref>. Rhodopsin is involved in visual signal transduction and the visual system in classic G protein-coupled receptor mechanisms<ref name="Article12">PMID:11891118</ref>. | + | '''Rhodopsin,''' a homodimeric protein, is a highly characterized [[G protein-coupled receptor]] found in membranous disks of the outer segments of rod and cone cells, though rhodopsin is more concentrated in rod cells which are sensitive to light but cannot discriminate colors. Rhodopsin is part of the superfamily of G protein-coupled receptors that mediate responses to visual, olfactory, hormonal, and neurotransmitter signals among others<ref name="Article1">PMID:20004206</ref>. Rhodopsin is involved in visual signal transduction and the visual system in classic G protein-coupled receptor mechanisms<ref name="Article12">PMID:11891118</ref>. |
| + | *'''Sodium pump rhodopsin''' transports Na+ and H+ depending on ionic conditions<ref>PMID:28749956</ref>. | ||
| + | *'''Chloride pump rhodopsin''' transports Cl- into cells<ref>PMID:33753488</ref>. | ||
| + | *'''Metarhodopsin II''' is a rhodopsin intermediate in which the retinal Schiff base is still intact but deprotonated<ref>PMID:21389988</ref>. | ||
| + | *'''Bathorhodopsin''' and '''Lumirhodopsin''' are rhodopsin complexes with metastable intermediates of retinal<ref>PMID:21341699</ref>,<ref>PMID:11425321</ref>. | ||
| + | *'''Bestrhodopsin''' is a rhodopsin-rhodopsin-bestrophin complex<ref>PMID:35710843</ref>. | ||
| + | *'''Cyanorhodopsin''' is a rhodopsin found in cyanobacteria<ref>PMID:33028840</ref>. | ||
| + | *For '''Halorhodopsin''', '''Archaerhodopsin''', '''Proteorhodopsin''', '''Xanthorhodopsin''' , '''Deltarhodopsin''' see [[Bacteriorhodopsin]]. | ||
| + | |||
| + | |||
| + | See also [[Transmembrane (cell surface) receptors]], [[Bacteriorhodopsin]] | ||
===G Protein-Coupled Receptors=== | ===G Protein-Coupled Receptors=== | ||
| - | Rhodopsin is a member of the superfamily of [[G protein-coupled receptor|G protein-coupled receptors]] that incorporate the activation of G proteins in their modulation of signaling and intracellular actions. Rhodopsin shares similar membrane topology with the members of the superfamily (Family A of the [[G protein-coupled receptor|G protein-coupled receptors]]) which include the seven transmembrane helices, an extracellular N terminus and cytoplasmic C terminus<ref name="Article20">PMID:15251227</ref>. The seven-helical pattern is found from archaebacteria (specifically studied is bacteriorhodopsin) to humans, both which share the same retinylidene chromophore as well <ref name="Article12"/>. Given that the the seven transmembrane domain of rhodopsin was the first solved crystal structure and that very few have been determined for other members of the G protein-coupled receptor superfamily, rhodopsin often serves as a reference for the structure and function relationship for other [[G protein-coupled receptor|G protein-coupled receptors]]<ref name="Article20"/><ref name="RecentOthers">PMID:20019124</ref>. Like most G protein-coupled receptors, the activated rhodopsin catalyzes uptake of GTP by the heterotrimeric G protein, in this case [http://en.wikipedia.org/wiki/Transducin transducin], which interacts with the cytoplasmic loops of the receptor<ref name="Article10">PMID:11698103</ref>. However, the covalent binding nature of rhodopsin to its retinal ligand is unlike most G protein-coupled receptors. Additionally, most other members of the superfamily use small ligands as inducers, whereas rhodopsin uses light as the inducer for activation<ref name="Article20"/>. | + | Rhodopsin is a member of the superfamily of [[G protein-coupled receptor|G protein-coupled receptors]] that incorporate the activation of G proteins in their modulation of signaling and intracellular actions. Rhodopsin shares similar membrane topology with the members of the superfamily (Family A of the [[G protein-coupled receptor|G protein-coupled receptors]]) which include the seven transmembrane helices, an extracellular N terminus and cytoplasmic C terminus<ref name="Article20">PMID:15251227</ref>. The seven-helical pattern is found from archaebacteria (specifically studied is bacteriorhodopsin) to humans, both which share the same retinylidene chromophore as well <ref name="Article12"/>. Given that the the seven transmembrane domain of rhodopsin was the first solved crystal structure and that very few have been determined for other members of the G protein-coupled receptor superfamily, rhodopsin often serves as a reference for the structure and function relationship for other [[G protein-coupled receptor|G protein-coupled receptors]]<ref name="Article20"/><ref name="RecentOthers">PMID:20019124</ref>. Like most G protein-coupled receptors, the activated rhodopsin catalyzes uptake of GTP by the heterotrimeric G protein, in this case [http://en.wikipedia.org/wiki/Transducin transducin] ([[Transducin]]), which interacts with the cytoplasmic loops of the receptor<ref name="Article10">PMID:11698103</ref>. However, the covalent binding nature of rhodopsin to its retinal ligand is unlike most G protein-coupled receptors. Additionally, most other members of the superfamily use small ligands as inducers, whereas rhodopsin uses light as the inducer for activation<ref name="Article20"/>. |
==Structure== | ==Structure== | ||
===Rhodopsin Architecture=== | ===Rhodopsin Architecture=== | ||
| - | Rhodopsin consists of seven mostly α-helical transmembrane domains (H1-H7) linked sequentially by extracellular and cytoplasmic loops (E1-E3 and C1-C3 respectively), with the extracellular amino-terminal tail and the cytoplasmic carboxyl-terminal tail<ref name="Article12"/>. Four of the helices are tilted and three of the helices are approximately perpendicular to the membrane plane<ref name="Article4">PMID:9199406</ref>. There is notable interaction between the four extracellular domains, but only a few associations are observed with the cytoplasmic domains<ref name="Article9">PMID:11343925</ref>. Helix 7 is close to being elongated around the Lysine 296 retinal attachment site, and also contains the residues Proline 291 and Proline 303, with Proline 303 being part of a conserved motif<ref name="Article9"/>. Near the retinal region, there is a <scene name=' | + | Rhodopsin consists of seven mostly α-helical transmembrane domains (H1-H7) linked sequentially by extracellular and cytoplasmic loops (E1-E3 and C1-C3 respectively), with the extracellular amino-terminal tail and the cytoplasmic carboxyl-terminal tail<ref name="Article12"/>. Four of the helices are tilted and three of the helices are approximately perpendicular to the membrane plane<ref name="Article4">PMID:9199406</ref>. There is notable interaction between the four extracellular domains, but only a few associations are observed with the cytoplasmic domains<ref name="Article9">PMID:11343925</ref>. Helix 7 is close to being elongated around the Lysine 296 retinal attachment site, and also contains the residues Proline 291 and Proline 303, with Proline 303 being part of a conserved motif<ref name="Article9"/>. Near the retinal region, there is a <scene name='40/400594/Cv/2'>β4 strand (Serine 186-Cysteine 187-Glycine 188-Isoleucine 189)</scene> within the Extracellular Helix 2 that runs almost parallel to the chromophore held in place and is stabilized by the essential conserved |
| - | <scene name=' | + | <scene name='40/400594/Cv/3'>disulfide bond between Cysteine 110 and Cysteine 187</scene>. This loop also potentially contacts the chromophore through Glutamine 181 and Tyrosine 191<ref name="Article12"/>. |
<scene name='Sandbox_173/Water_molecules/1'>Water molecules</scene> are observed to be located in the extracellular domains of rhodopsin; specifically, the water molecules around the second extracellular loop between Helix 4 and 5 solvate the loop when the loop interacts with the retinal chromophore and possibly contribute to its flexibility should rearrangement occur<ref name="ReferenceArticle">PMID:15327956</ref>. | <scene name='Sandbox_173/Water_molecules/1'>Water molecules</scene> are observed to be located in the extracellular domains of rhodopsin; specifically, the water molecules around the second extracellular loop between Helix 4 and 5 solvate the loop when the loop interacts with the retinal chromophore and possibly contribute to its flexibility should rearrangement occur<ref name="ReferenceArticle">PMID:15327956</ref>. | ||
There is the presence of a cationic amphipathic Helix 8, known as the fourth cytoplasmic loop, that spans from <scene name='Sandbox_173/Helix_8/1'>Asparagine 310 to Cysteine 323</scene> and is formed from the C-terminal tail anchoring to the membrane by | There is the presence of a cationic amphipathic Helix 8, known as the fourth cytoplasmic loop, that spans from <scene name='Sandbox_173/Helix_8/1'>Asparagine 310 to Cysteine 323</scene> and is formed from the C-terminal tail anchoring to the membrane by | ||
| - | <scene name=' | + | <scene name='40/400594/Cv/4'>Cysteine 322 and Cysteine 323</scene>, which are <scene name='40/400594/Cv/5'>palmitoylated</scene>. This helix runs approximately parallel to the cytoplasmic surface and is involved in Gtγ binding<ref name="Article9"/>, as well as the modulation of rhodopsin-transducin interactions and rhodopsin-phospholipid interactions<ref name="Article12"/>. |
A metal zinc ion bridge chelated by histidine side-chains and connected to the cytoplasmic ends of Helix 3 and 6 is observed to prevent receptor activation. This perhaps indicates that separation of these cytoplasmic ends would contribute to rhodopsin activation<ref name="Article10"/>. | A metal zinc ion bridge chelated by histidine side-chains and connected to the cytoplasmic ends of Helix 3 and 6 is observed to prevent receptor activation. This perhaps indicates that separation of these cytoplasmic ends would contribute to rhodopsin activation<ref name="Article10"/>. | ||
| Line 22: | Line 33: | ||
===Retinal Chromophore of Rhodopsin=== | ===Retinal Chromophore of Rhodopsin=== | ||
| - | Rhodopsin consists of an opsin [http://en.wikipedia.org/wiki/Apoprotein apoprotein] and a <scene name=' | + | Rhodopsin consists of an opsin [http://en.wikipedia.org/wiki/Apoprotein apoprotein] and a <scene name='40/400594/Cv/7'>11-cis retinylidene chromophore</scene> in its active site. Rhodopsin is bound covalently to the 11-''cis'' retinal, the chromophore or "ligand," (shown in <span style="color:yellow;background-color:black;font-weight:bold;">yellow</span>) and this retinal is found in deeply in the core of the helices, in a hydrophobic site, parallel to the lipid bilayer<ref name="Article19">PMID:16051215</ref>. Comparatively, it is situated more towards the extracellular planes of the membrane bilayer <ref name="Article12"/>. The retinal is attached in the active site of rhodopsin through a protonated Schiff base (an N-substituted imine) bond to the ε-amino group of Lysine 296 residue (shown in <span style="color:lime;background-color:black;font-weight:bold;">green</span>) on the C-terminal Helix 7, with this linkage creating a positive charge on the chromophore <ref name="Article4"/>. The protonated Schiff base of rhodopsin is stabilized through <scene name='40/400594/Cv/8'>Glutamine 113</scene> residue electrostatic interaction with the counterion, holding the inactive rhodopsin in its state<ref name="Article20"/>. |
As this ligand is bound in the 12-s-''trans'' conformation, there arises the non-bonding interactions between the C-13 methyl group and C-10 hydrogen that contribute to non-planarity. This leads to the ability of the chromophore polyene tail to undergo fast photoisomerization around the C-11=C-12 double bond during light-induced activation<ref name="Article2">PMID:16962138</ref>. Also, it is found that the C-11=C-12 double bond is pre-twisted in the ground state of rhodopsin, which is partly attributed to the C20 methyl group attached to C13 through interaction with Tryptophan 265. This pre-twist may give insight on the features of isomerization about this bond upon light activation<ref name="ReferenceArticle"/>. | As this ligand is bound in the 12-s-''trans'' conformation, there arises the non-bonding interactions between the C-13 methyl group and C-10 hydrogen that contribute to non-planarity. This leads to the ability of the chromophore polyene tail to undergo fast photoisomerization around the C-11=C-12 double bond during light-induced activation<ref name="Article2">PMID:16962138</ref>. Also, it is found that the C-11=C-12 double bond is pre-twisted in the ground state of rhodopsin, which is partly attributed to the C20 methyl group attached to C13 through interaction with Tryptophan 265. This pre-twist may give insight on the features of isomerization about this bond upon light activation<ref name="ReferenceArticle"/>. | ||
Somewhat enclosing this chromophore is a retinal binding pocket partially formed by the N-terminal domain overlaying the extracellular turns including the second extracellular loop, which folds into the molecular center<ref name="Article6">PMID:18692154</ref>. | Somewhat enclosing this chromophore is a retinal binding pocket partially formed by the N-terminal domain overlaying the extracellular turns including the second extracellular loop, which folds into the molecular center<ref name="Article6">PMID:18692154</ref>. | ||
| - | |||
==Function== | ==Function== | ||
| Line 38: | Line 48: | ||
====Formation of the Metarhodopsin II State==== | ====Formation of the Metarhodopsin II State==== | ||
| - | Rhodopsin forms to Metarhodopsin II, the intermediate signaling state where interaction occurs with the G protein. This millisecond process is accompanied by movement in the helices, uptake of protons in the cytoplasm, and the breakage of the salt bridge between Glutamine 113 and the protonated Schiff base. The Schiff base deprotonates and the proton is transferred to the Glutamine 113 counterion, destabilizing the ground state <ref name="Article9"/>. As well, this Metarhodopsin II formation may be dependent on the protonation too of the conserved <scene name=' | + | Rhodopsin forms to Metarhodopsin II, the intermediate signaling state where interaction occurs with the G protein. This millisecond process is accompanied by movement in the helices, uptake of protons in the cytoplasm, and the breakage of the salt bridge between Glutamine 113 and the protonated Schiff base. The Schiff base deprotonates and the proton is transferred to the Glutamine 113 counterion, destabilizing the ground state <ref name="Article9"/>. As well, this Metarhodopsin II formation may be dependent on the protonation too of the conserved <scene name='40/400594/Cv/9'>Glutamine 134 that forms a salt bridge with Arginine 135</scene>, thus destabilizing the constraint on Arginine 135<ref name="Article9"/>. |
There is positive enthalpy associated with the formation of Metarhodopsin II. This formation of the active state, also linked with the increase in entropy, is suggested to release the constraints in the helices and expose the cytoplasmic binding sites<ref name="Article9"/>. An important part of this process includes the 9-methyl group of retinal, which is suggested to provide a scaffold for proton transfers essential for the formation of the active state<ref name="Article9"/>. | There is positive enthalpy associated with the formation of Metarhodopsin II. This formation of the active state, also linked with the increase in entropy, is suggested to release the constraints in the helices and expose the cytoplasmic binding sites<ref name="Article9"/>. An important part of this process includes the 9-methyl group of retinal, which is suggested to provide a scaffold for proton transfers essential for the formation of the active state<ref name="Article9"/>. | ||
| Line 53: | Line 63: | ||
====Phosphorylation and Deactivation of Rhodopsin==== | ====Phosphorylation and Deactivation of Rhodopsin==== | ||
| - | [http://en.wikipedia.org/wiki/Rhodopsin_kinase Rhodopsin kinase] phosphorylates rhodopsin and [http://en.wikipedia.org/wiki/Arrestin arrestin] binds to the phosphorylated domain of rhodopsin, preventing further signal transduction from Metarhodopsin II of activated rhodopsin and transducin<ref name="Article3"/>. It phosphorylates both Metarhodopsin II and cone opsins. The majority of the phosphorylation sites are in the cytoplasmic C-terminal region of rhodopsin with seven hydroxy-amino acids. The most favoured amino acids are <scene name=' | + | [http://en.wikipedia.org/wiki/Rhodopsin_kinase Rhodopsin kinase] phosphorylates rhodopsin and [http://en.wikipedia.org/wiki/Arrestin arrestin] binds to the phosphorylated domain of rhodopsin, preventing further signal transduction from Metarhodopsin II of activated rhodopsin and transducin<ref name="Article3"/>. It phosphorylates both Metarhodopsin II and cone opsins. The majority of the phosphorylation sites are in the cytoplasmic C-terminal region of rhodopsin with seven hydroxy-amino acids. The most favoured amino acids are <scene name='40/400594/Cv/10'>Serine 338, Serine 343, Serine 334, Threonine 335 and Threonine 336</scene><ref name="Article7">PMID:9667002</ref>, and these residues form an arrangement in rhodopsin that do not appear to be exposed to the solvent. Interactions with the C-terminal tail and a portion of the Cytoplasmic loop 3 appear to be broken for the phosphorylation of the hydroxyl groups<ref name="Article9"/>. For the next cycle of activation of rhodopsin, rhodopsin has to be dephosphorylated, and have the all-''trans'' retinal replaced with the 11-''cis'' retinal<ref name="Article19"/>. |
Altogether, the different states of rhodopsin which include the short-lived, photo-rhodopsin, batho-rhodopsin, and lumi-rhodopsin, and longer-lived meta-rhodopsins give information about the structural status of the molecule during activation<ref name="Article9"/>. | Altogether, the different states of rhodopsin which include the short-lived, photo-rhodopsin, batho-rhodopsin, and lumi-rhodopsin, and longer-lived meta-rhodopsins give information about the structural status of the molecule during activation<ref name="Article9"/>. | ||
| + | See also [[Rhodopsin Structure and Function]]. | ||
==Opsin== | ==Opsin== | ||
===Topology Overview=== | ===Topology Overview=== | ||
| - | The overall dimeric structure of opsin is similar to rhodopsin, with seven transmembrane helices linked by three extracellular loops and three cytoplasmic loops and a cytoplasmic Helix 8. The small differences between the topology of the two proteins include a short helical turn in the cytoplasmic loop 1 in opsin, 1.5-2.5 helical turns longer in Helix 5 for opsin in comparison to rhodopsin, and a large outward tilt of Helix 6 of opsin<ref name="ArticleOpsin2">PMID:18563085</ref>. Also, in contrast to rhodopsin, opsin has two openings of the retinal-binding pocket; one of the openings is between Helix 1 and Helix 7, and the other opening is between the extracellular ends of Helix 5 and 6. This opening is formed by the residues <scene name=' | + | The overall dimeric structure of opsin is similar to rhodopsin, with seven transmembrane helices linked by three extracellular loops and three cytoplasmic loops and a cytoplasmic Helix 8. The small differences between the topology of the two proteins include a short helical turn in the cytoplasmic loop 1 in opsin, 1.5-2.5 helical turns longer in Helix 5 for opsin in comparison to rhodopsin, and a large outward tilt of Helix 6 of opsin<ref name="ArticleOpsin2">PMID:18563085</ref>. Also, in contrast to rhodopsin, opsin has two openings of the retinal-binding pocket; one of the openings is between Helix 1 and Helix 7, and the other opening is between the extracellular ends of Helix 5 and 6. This opening is formed by the residues <scene name='40/400594/Cv/11'>Isoleucine 205 and Phenylalanine 208 in Helix 5, and by the residues Phenylalanine 273 and Phenylalanine 276 in Helix 6</scene><ref name="ArticleOpsin2"/>. The two openings suggest different sites of retinal entrance and exit in retinal channeling<ref name="ArticleOpsin2"/>. |
===Activity=== | ===Activity=== | ||
| Line 67: | Line 78: | ||
===Colour Vision=== | ===Colour Vision=== | ||
Opsins are also photoreceptor proteins and are concentrated in cone cells, cells that are less sensitive to light but can discriminate colours. Opsins are slightly different light receptors than rhodopsin in that they can detect light from different spectrums and distinguish between their wavelengths. The ability to differentiate between colours is related to the three types of cone cells, each using one of the three related opsin photoreceptors<ref name="Textbook"/>. | Opsins are also photoreceptor proteins and are concentrated in cone cells, cells that are less sensitive to light but can discriminate colours. Opsins are slightly different light receptors than rhodopsin in that they can detect light from different spectrums and distinguish between their wavelengths. The ability to differentiate between colours is related to the three types of cone cells, each using one of the three related opsin photoreceptors<ref name="Textbook"/>. | ||
| - | |||
† PDB structure used in this section: [[3cap]] | † PDB structure used in this section: [[3cap]] | ||
| - | __NOTOC__ | ||
__NOEDITSECTION__ | __NOEDITSECTION__ | ||
| - | </StructureSection> | ||
==3D structures of rhodopsin== | ==3D structures of rhodopsin== | ||
| + | [[Rhodopsin 3D structures]] | ||
| - | + | </StructureSection> | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
==References== | ==References== | ||
| Line 113: | Line 93: | ||
==See Also== | ==See Also== | ||
| + | * [[Bacteriorhodopsin]] | ||
*[[G protein-coupled receptor|G protein-coupled receptors]] | *[[G protein-coupled receptor|G protein-coupled receptors]] | ||
*[[Membrane proteins]] | *[[Membrane proteins]] | ||
| + | *[[Receptor]] | ||
| + | *[[Transmembrane (cell surface) receptors]] | ||
[[Category:Topic Page]] | [[Category:Topic Page]] | ||
| Line 121: | Line 104: | ||
[[Category: Photoreceptor]] | [[Category: Photoreceptor]] | ||
[[Category: Retinal protein]] | [[Category: Retinal protein]] | ||
| - | __NOTOC__ | ||
Current revision
| |||||||||||

References
- ↑ Hornak V, Ahuja S, Eilers M, Goncalves JA, Sheves M, Reeves PJ, Smith SO. Light activation of rhodopsin: insights from molecular dynamics simulations guided by solid-state NMR distance restraints. J Mol Biol. 2010 Feb 26;396(3):510-27. Epub 2009 Dec 11. PMID:20004206 doi:10.1016/j.jmb.2009.12.003
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 Sakmar TP. Structure of rhodopsin and the superfamily of seven-helical receptors: the same and not the same. Curr Opin Cell Biol. 2002 Apr;14(2):189-95. PMID:11891118
- ↑ Tsunoda SP, Prigge M, Abe-Yoshizumi R, Inoue K, Kozaki Y, Ishizuka T, Yawo H, Yizhar O, Kandori H. Functional characterization of sodium-pumping rhodopsins with different pumping properties. PLoS One. 2017 Jul 27;12(7):e0179232. PMID:28749956 doi:10.1371/journal.pone.0179232
- ↑ Yun JH, Li X, Yue J, Park JH, Jin Z, Li C, Hu H, Shi Y, Pandey S, Carbajo S, Boutet S, Hunter MS, Liang M, Sierra RG, Lane TJ, Zhou L, Weierstall U, Zatsepin NA, Ohki M, Tame JRH, Park SY, Spence JCH, Zhang W, Schmidt M, Lee W, Liu H. Early-stage dynamics of chloride ion-pumping rhodopsin revealed by a femtosecond X-ray laser. Proc Natl Acad Sci U S A. 2021 Mar 30;118(13). pii: 2020486118. doi:, 10.1073/pnas.2020486118. PMID:33753488 doi:http://dx.doi.org/10.1073/pnas.2020486118
- ↑ Choe HW, Kim YJ, Park JH, Morizumi T, Pai EF, Krauss N, Hofmann KP, Scheerer P, Ernst OP. Crystal structure of metarhodopsin II. Nature. 2011 Mar 9. PMID:21389988 doi:10.1038/nature09789
- ↑ Schapiro I, Ryazantsev MN, Frutos LM, Ferré N, Lindh R, Olivucci M. The ultrafast photoisomerizations of rhodopsin and bathorhodopsin are modulated by bond length alternation and HOOP driven electronic effects. J Am Chem Soc. 2011 Mar 16;133(10):3354-64. PMID:21341699 doi:10.1021/ja1056196
- ↑ Pan D, Mathies RA. Chromophore structure in lumirhodopsin and metarhodopsin I by time-resolved resonance Raman microchip spectroscopy. Biochemistry. 2001 Jul 3;40(26):7929-36. PMID:11425321 doi:10.1021/bi010670x
- ↑ Rozenberg A, Kaczmarczyk I, Matzov D, Vierock J, Nagata T, Sugiura M, Katayama K, Kawasaki Y, Konno M, Nagasaka Y, Aoyama M, Das I, Pahima E, Church J, Adam S, Borin VA, Chazan A, Augustin S, Wietek J, Dine J, Peleg Y, Kawanabe A, Fujiwara Y, Yizhar O, Sheves M, Schapiro I, Furutani Y, Kandori H, Inoue K, Hegemann P, Béjà O, Shalev-Benami M. Rhodopsin-bestrophin fusion proteins from unicellular algae form gigantic pentameric ion channels. Nat Struct Mol Biol. 2022 Jun;29(6):592-603. PMID:35710843 doi:10.1038/s41594-022-00783-x
- ↑ Hasegawa M, Hosaka T, Kojima K, Nishimura Y, Nakajima Y, Kimura-Someya T, Shirouzu M, Sudo Y, Yoshizawa S. A unique clade of light-driven proton-pumping rhodopsins evolved in the cyanobacterial lineage. Sci Rep. 2020 Oct 7;10(1):16752. PMID:33028840 doi:10.1038/s41598-020-73606-y
- ↑ 10.0 10.1 10.2 10.3 Kristiansen K. Molecular mechanisms of ligand binding, signaling, and regulation within the superfamily of G-protein-coupled receptors: molecular modeling and mutagenesis approaches to receptor structure and function. Pharmacol Ther. 2004 Jul;103(1):21-80. PMID:15251227 doi:10.1016/j.pharmthera.2004.05.002
- ↑ Millar RP, Newton CL. The year in G protein-coupled receptor research. Mol Endocrinol. 2010 Jan;24(1):261-74. Epub 2009 Dec 17. PMID:20019124 doi:10.1210/me.2009-0473
- ↑ 12.0 12.1 12.2 Meng EC, Bourne HR. Receptor activation: what does the rhodopsin structure tell us? Trends Pharmacol Sci. 2001 Nov;22(11):587-93. PMID:11698103
- ↑ 13.0 13.1 Shieh T, Han M, Sakmar TP, Smith SO. The steric trigger in rhodopsin activation. J Mol Biol. 1997 Jun 13;269(3):373-84. PMID:9199406 doi:10.1006/jmbi.1997.1035
- ↑ 14.0 14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 Okada T, Ernst OP, Palczewski K, Hofmann KP. Activation of rhodopsin: new insights from structural and biochemical studies. Trends Biochem Sci. 2001 May;26(5):318-24. PMID:11343925
- ↑ 15.0 15.1 Okada T, Sugihara M, Bondar AN, Elstner M, Entel P, Buss V. The retinal conformation and its environment in rhodopsin in light of a new 2.2 A crystal structure. J Mol Biol. 2004 Sep 10;342(2):571-83. PMID:15327956 doi:10.1016/j.jmb.2004.07.044
- ↑ 16.0 16.1 Janz JM, Farrens DL. Assessing structural elements that influence Schiff base stability: mutants E113Q and D190N destabilize rhodopsin through different mechanisms. Vision Res. 2003 Dec;43(28):2991-3002. PMID:14611935
- ↑ 17.0 17.1 17.2 Kisselev OG. Focus on molecules: rhodopsin. Exp Eye Res. 2005 Oct;81(4):366-7. PMID:16051215 doi:10.1016/j.exer.2005.06.018
- ↑ 18.0 18.1 18.2 Verhoeven MA, Bovee-Geurts PH, de Groot HJ, Lugtenburg J, DeGrip WJ. Methyl substituents at the 11 or 12 position of retinal profoundly and differentially affect photochemistry and signalling activity of rhodopsin. J Mol Biol. 2006 Oct 13;363(1):98-113. Epub 2006 Jul 28. PMID:16962138 doi:10.1016/j.jmb.2006.07.039
- ↑ 19.0 19.1 19.2 19.3 Morris MB, Dastmalchi S, Church WB. Rhodopsin: structure, signal transduction and oligomerisation. Int J Biochem Cell Biol. 2009 Apr;41(4):721-4. Epub 2008 Aug 3. PMID:18692154 doi:10.1016/j.biocel.2008.04.025
- ↑ 20.0 20.1 20.2 20.3 20.4 Nelson, D., and Cox, M. Lehninger Principles of Biochemistry. 2008. 5th edition. W. H. Freeman and Company, New York, New York, USA. pp. 462-465.
- ↑ Hurley JB, Spencer M, Niemi GA. Rhodopsin phosphorylation and its role in photoreceptor function. Vision Res. 1998 May;38(10):1341-52. PMID:9667002
- ↑ 22.0 22.1 22.2 Park JH, Scheerer P, Hofmann KP, Choe HW, Ernst OP. Crystal structure of the ligand-free G-protein-coupled receptor opsin. Nature. 2008 Jul 10;454(7201):183-7. Epub 2008 Jun 18. PMID:18563085 doi:10.1038/nature07063
- ↑ 23.0 23.1 Surya A, Knox BE. Enhancement of opsin activity by all-trans-retinal. Exp Eye Res. 1998 May;66(5):599-603. PMID:9628807 doi:10.1006/exer.1997.0453
See Also
- Bacteriorhodopsin
- G protein-coupled receptors
- Membrane proteins
- Receptor
- Transmembrane (cell surface) receptors
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, Alexander Berchansky, Wayne Decatur, Jaime Prilusky, Joel L. Sussman, Cinting Lim
