From Proteopedia
proteopedia linkproteopedia link
The FpgNei Protein Superfamily
Functional Units
| G. Stereothermophilus Fpg
| E. Coli Nei
|
|
|
|
| Functional Cluster | Variant 1 | Variant 2 | Fpg1 | Fpg2 | Plant | Neil1 | Neil2 | Neil3 | Proteo | Actino1 | Actino2 | MimiVirus
|
| Support for perfectly conserved Asn168
|
|
| Y | Y | N | N | N | N | N | N | N | N
|
| Stability of catalytic helix
|
|
| Y | Y | Y | N | N | N | N | N | N | N
|
| Stability of intercalation loop
|
|
| Y | Y | Y | N | N | N | N | N | N | Y
|
| Intercalation loop
|
|
| Y | Y | Y | N | N | N | N | N | N | Y
|
| Zinc Finger
|
|
| Y | Y | N | N | Y | Y | Y | Y | Y | N
|
| Recognition complex
|
| none
| Y | N | N | N | N | N | N | N | N | N
|
| Neil1-specific: Support for Lys60
|
|
| Y | Y | Y | N | Y | Y | Y | Y | Y | Y
|
| PlantFungi-specific: R254 DNA binding
|
| different in plants
| Y | Y | N | Y | Y | Y | N | Y | Y | Y
|
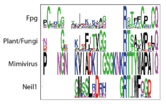
Functional Cluster: Stability of perfectly conserved Asn168
 WebLogo for the first functional unit, note the covariation between lysine nad arginine, which support the crucial Asn174 |
Functional Cluster: Stability of catalytic helix
 The LER triad interact in stabilize the catalytic helix (Pro2 and Glu3), it is not clear how this is achieved in the other subfamilies |
Functional Cluster: Stability of intercalation loop
The intercalation loop inserts into the DNA when the base is everted. This structure, as well as a group of amino acids that support it exhibit high conservation in Fpg, AY, plants and Neil1, but not on the rest. Note that the amino acid composition suggests that mimivirus has this structure.
|
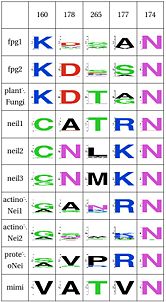
Functional Cluster: Stability of key Gly59 and Lys60
 Notice the covariation in Neil1 and the rest with positions 134 and 137 and 170 |
Functional Cluster: Intercalation Loop
The intercalation loop inserts into the space left by the excised base.
The residue E2 and E6 have been mutated, with the first one inactivating the protein and the second one having no major effect [1].
|
Functional Cluster: Zinc/zincless finger
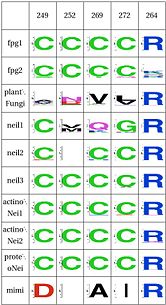 The Zinc Finger helps as a support of the key R264 residue |
Functional Cluster: Recognition Complex
This complex is key in recognizing a damaged guanine[5]
|
Functional Cluster: DNA binding Tyrosine
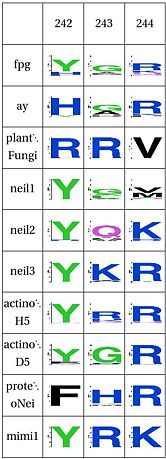 Tyrosine that binds to DNA, the plant has a different structure alltogether |
Zinc/zincless finger
Tyrosine that binds to DNA, the plant has a different structure alltogether
Evolution
The FpgNei evolution has not been easy to resolve [6], especially in the deeper branches. Assuming that functional clusters evolve more slowly than individual residues, we can use this as phylogenetic characters to 1) draw the most parsimonious evolution of the superfamily as dictated by these functional clusters 2) examine how these clusters have evolved and how this might have influenced the evolution of FpgNei.
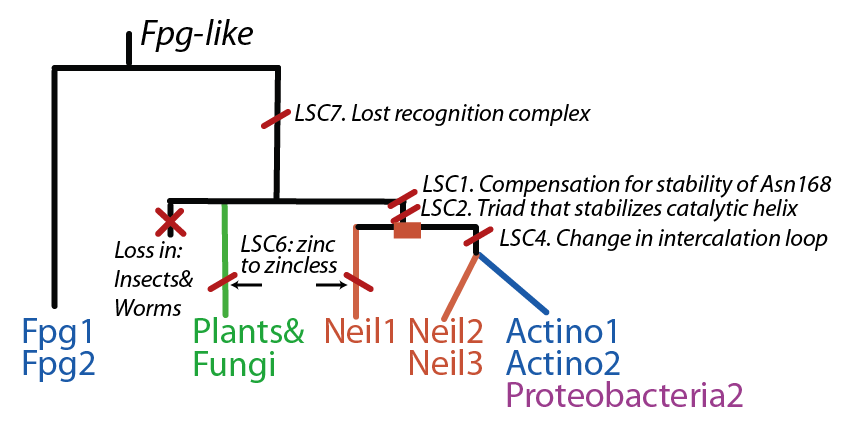
Site-directed mutants
| Name | Organism/Structure | Effect | Reference
|
| | Escherichia coli (1k3w) | inactive | [7][8]
|
| | Escherichia coli (1k82) | reduced glygosylase, less efficient in Schiff-base complex. | [9][10]
|
| | Escherichia coli (1k3w) | inactive, although can cleave AP sites | [11]
|
| | Escherichia coli (1k3w) | Inactive but can still make Schiff Base. | [12]
|
| | Escherichia coli (1k3w) | Decrease glycosylase, lyase ok. | [13]
|
| | Escherichia coli (1k3w) | Decrease glycosylase, lyase ok. | [14]
|
| | Homo Sapiens (1tdh) | Low activity | [15]
|
| | E. Coli (1k82) | No binding nor cleavage | [16]
|
| | E. Coli (1k82) | No binding nor cleavage. No zinc, no altered secondary structure | [17]
|
| | E. Coli (1k82) | No lyase nor glycosylase | [18]
|
|
|
References
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Fromme JC, Verdine GL. DNA lesion recognition by the bacterial repair enzyme MutM. J Biol Chem. 2003 Dec 19;278(51):51543-8. Epub 2003 Oct 1. PMID:14525999 doi:10.1074/jbc.M307768200
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Golan G, Zharkov DO, Feinberg H, Fernandes AS, Zaika EI, Kycia JH, Grollman AP, Shoham G. Structure of the uncomplexed DNA repair enzyme endonuclease VIII indicates significant interdomain flexibility. Nucleic Acids Res. 2005 Sep 6;33(15):5006-16. Print 2005. PMID:16145054 doi:http://dx.doi.org/33/15/5006
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Sidorkina OM, Laval J. Role of lysine-57 in the catalytic activities of Escherichia coli formamidopyrimidine-DNA glycosylase (Fpg protein). Nucleic Acids Res. 1998 Dec 1;26(23):5351-7. PMID:9826758
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Golan G, Zharkov DO, Feinberg H, Fernandes AS, Zaika EI, Kycia JH, Grollman AP, Shoham G. Structure of the uncomplexed DNA repair enzyme endonuclease VIII indicates significant interdomain flexibility. Nucleic Acids Res. 2005 Sep 6;33(15):5006-16. Print 2005. PMID:16145054 doi:http://dx.doi.org/33/15/5006
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Shinmura K, Tao H, Goto M, Igarashi H, Taniguchi T, Maekawa M, Takezaki T, Sugimura H. Inactivating mutations of the human base excision repair gene NEIL1 in gastric cancer. Carcinogenesis. 2004 Dec;25(12):2311-7. Epub 2004 Aug 19. PMID:15319300 doi:10.1093/carcin/bgh267
- ↑ Tchou J, Michaels ML, Miller JH, Grollman AP. Function of the zinc finger in Escherichia coli Fpg protein. J Biol Chem. 1993 Dec 15;268(35):26738-44. PMID:8253809
- ↑ Tchou J, Michaels ML, Miller JH, Grollman AP. Function of the zinc finger in Escherichia coli Fpg protein. J Biol Chem. 1993 Dec 15;268(35):26738-44. PMID:8253809
- ↑ O'Connor TR, Graves RJ, de Murcia G, Castaing B, Laval J. Fpg protein of Escherichia coli is a zinc finger protein whose cysteine residues have a structural and/or functional role. J Biol Chem. 1993 Apr 25;268(12):9063-70. PMID:8473347








{kind=link}
{kind=link}