This is a default text for your page Vinícius M. Neto/Sandbox 1. Click above on edit this page to modify. Be careful with the < and > signs.
You may include any references to papers as in: the use of JSmol in Proteopedia [1] or to the article describing Jmol [2] to the rescue.
Introduction
Silk fibers from Bombyx mori (silkworm) have been utilized by mankind since ancient times due to their remarkable mechanical properties and comfort when woven into fabrics, with the earliest records dating to around 2700 BC [3]. They are thermally comfortable, elastic, strong, and soft—properties that make silk highly sought after as a luxury fabric for garments. Silk is also biocompatible, making it applicable as a medical biomaterial for sutures, drug delivery systems, and scaffolds, where it plays a vital role in tissue regeneration[4].
Fibroins are fibrous structural proteins composed of multiple subunits that assemble into high-strength materials like silk and byssal threads. The exact composition varies by organism but typically features core fibroin filaments bundled within a sericin coating. These core filaments consist of three key components: a heavy chain (FibH), a light chain (FibL), and glycoproteins, all stabilized by disulfide bonds. The N-terminal domain of FibH (FibNT) plays a crucial role in the pH-dependent assembly of fibroin molecules during fiber formation [5].
Silk fibroin transitions between two key structural states: Silk I, the soluble pre-spun form stored in silk glands, consists of type II β-turns that prevent premature crystallization, while Silk II, the spun fiber's insoluble crystalline form, is dominated by mechanically robust antiparallel β-sheets. The N-terminal domain of fibroin plays a critical regulatory role—its pH-sensitive structure (rich in acidic residues) helps maintain Silk I's solubility in the gland but triggers the shift to Silk II during spinning by undergoing conformational changes that nucleate β-sheet formation. This pH-driven mechanism, combined with shear forces and dehydration, ensures the precise, irreversible transition from Silk I's storage form to Silk II's functional fiber, optimizing silk's strength and biological utility.
Function
Basic structure
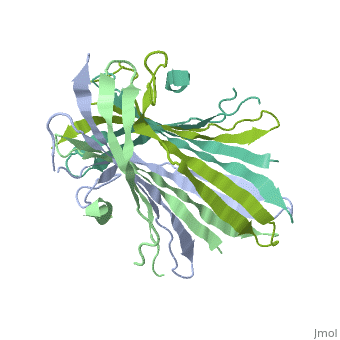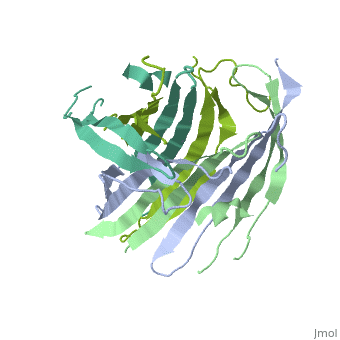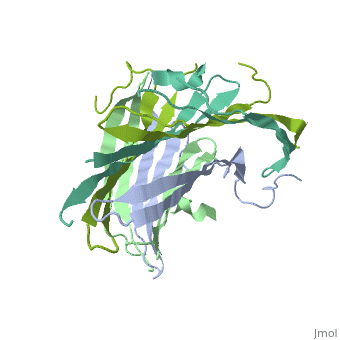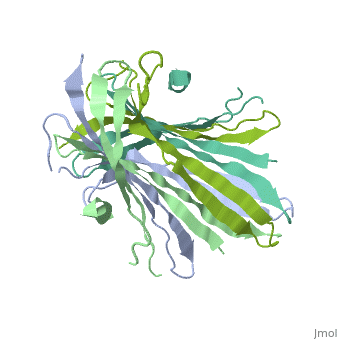
The N-terminal domain of the fibroin heavy chain (FibNT 3UA0) is a homo-tetramer composed of 268 residues, most of which are hydrophilic ( and ). FibNT's is a homodimer with eight alternating β- and a disordered (Gly109-Ser126). Its (A and B) are nearly identical except for the (Phe26-Val35):
- Chain A: Adopts a loop conformation.
- Chain B: Forms a short α-helix.
The FibNT homodimer exhibits the following : β1A–β2A–β4B–β3B–β3A–β4A–β2B–β1B, where there are two β-hairpins (Thr36–Asn65 and Glu78–Ser107) connected with two type I β-turns (Asp49–Gly52 and Asp89–Gly92). The entire assembly is stabilized by an extensive network of between adjacent β-strands.
pH-Dependent Structural Transition
The structure of fibroin is highly pH-dependent. During the natural silk-spinning process, the fibroin solution experiences a steep pH gradient along the silk gland (from anterior to posterior), which triggers the gelation of condensed fibroin. Specifically, the N-terminal domain (FibNT) remains in a disordered random-coil state at neutral pH, preventing premature β-sheet formation. Only when the pH drops to approximately 6.0 does FibNT undergo a cooperative structural transition, adopting the stable β-sheet conformation essential for fiber assembly.
Interactions between acidic residues in FibNT are critical for pH-sensitive behavior. Near the transition point (pH ~6.0), some residues exhibit up-shifted pKa values, allowing them to remain ionized at neutral pH. This sustained negative charge creates electrostatic repulsion, actively preventing premature folding and β-sheet assembly. For example, at higher pH, —such as those between Glu56–Asp44 and Asp100–Glu98—are disrupted, destabilizing β-sheet conformations until protonation occurs at lower pH.