Hepatocyte growth factor receptor
From Proteopedia
(Difference between revisions)
| (26 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
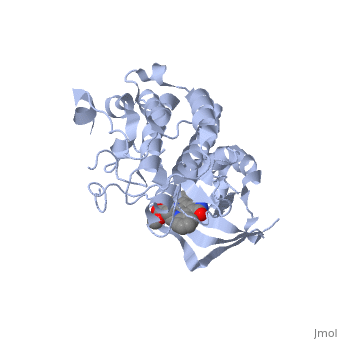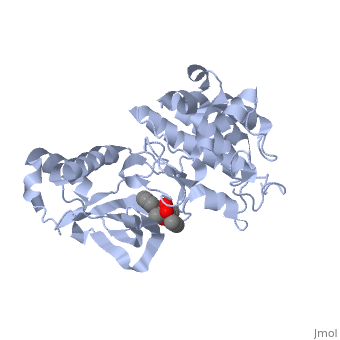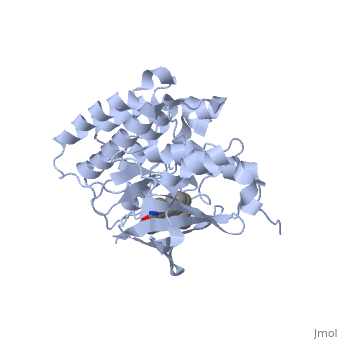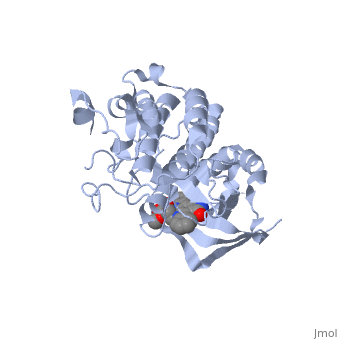
| + | <StructureSection load='1r0p' size='350' side='right' caption='Hepatocyte Growth Factor Receptor Tyrosine Kinase in complex with the inhibitor staurosporine analog K-252a (PDB entry [[1r0p]])' scene=''> | ||
=== Hepatocyte Growth Factor Receptor c-Met === | === Hepatocyte Growth Factor Receptor c-Met === | ||
| - | |||
| - | <StructureSection load='1r0p' size='350' side='right' caption='Hepatocyte Growth Factor Receptor Tyrosine Kinase (PDB entry [[1r0p]])' scene=''> | ||
==Introduction== | ==Introduction== | ||
| - | The Hepatocyte Growth Factor Receptor plays a large role in embryonic development, as its activation leads to events such as cell growth, motility and invasion. This receptor is a Tyrosine Kinase, and is one of the most well studied RTKs, as mutations in the c-met proto-oncogene can lead to the formation of tumors. The ligand for this receptor is Hepatocyte growth factor/scatter factor (HGF/SF), and upon binding of this ligand, the receptor becomes auto phosphorylated, causing downstream signaling events such as cell growth. <ref>PMID: 14559966</ref> C-Met is an αβ heterodimer with extracellular and intracellular domains. These two domains are disulfide linked together. <ref>PMID: 14559966</ref> This particular structure is one of a mutated hepatocyte growth factor tyrosine kinase domain, which is part of the intracellular β subunit. <ref>http://en.wikipedia.org/wiki/C-Met</ref> | + | The '''Hepatocyte Growth Factor Receptor''' or '''tyrosine-protein kinase Met''' or '''c-Met''' plays a large role in embryonic development, as its activation leads to events such as cell growth, motility and invasion. This receptor is a Tyrosine Kinase, and is one of the most well studied [[Receptor tyrosine kinases|RTKs]], as mutations in the c-met proto-oncogene can lead to the formation of tumors. The ligand for this receptor is Hepatocyte growth factor/scatter factor (HGF/SF), and upon binding of this ligand, the receptor becomes auto phosphorylated, causing downstream signaling events such as cell growth. <ref>PMID: 14559966</ref> C-Met is an αβ heterodimer with extracellular and intracellular domains. These two domains are disulfide linked together. <ref>PMID: 14559966</ref> This particular structure is one of a mutated hepatocyte growth factor tyrosine kinase domain, which is part of the intracellular β subunit.<ref>http://en.wikipedia.org/wiki/C-Met</ref>. See also [[Growth factors]] and [[Kinase-linked, enzyme-linked and related receptors]]. |
==Structure== | ==Structure== | ||
| Line 12: | Line 11: | ||
This receptor follows the typical structure of a protein kinase, with a bilobal structure. The N-terminal contains <scene name='Hepatocyte_growth_factor_receptor/Beta_sheets/1'>β-sheets</scene> and is linked through a hinge to the C lobe, which is full of α helices. This particular kinase domain is very similar to the domains of the insulin receptor kinase and fibroblast growth factor receptor kinase.<ref>PMID: 14559966</ref> | This receptor follows the typical structure of a protein kinase, with a bilobal structure. The N-terminal contains <scene name='Hepatocyte_growth_factor_receptor/Beta_sheets/1'>β-sheets</scene> and is linked through a hinge to the C lobe, which is full of α helices. This particular kinase domain is very similar to the domains of the insulin receptor kinase and fibroblast growth factor receptor kinase.<ref>PMID: 14559966</ref> | ||
| - | =Helices= | + | ==Helices== |
| - | This structure is made up of many α helical structures that move in the transformation from inactive to active kinase. Some of these helices are conserved in many different tyrosine kinases. C-met does show a divergence from other tyrosine kinases (such as IRK and FGFRK) in the helix formed at the N-terminus, before the core kinase domain, in residues <scene name='Hepatocyte_growth_factor_receptor/1060-1069/1'>1060-1069</scene>. <ref>PMID: 14559966</ref> The αA is in contact with αC and so causes αC to be in a slightly different orientation than in FGFRK and IRK. Residues Leu-1062, Val-1066, and Val-1069 of αA <scene name='Hepatocyte_growth_factor_receptor/A_and_c_intercalating/1'>intercalate</scene> with with residues Leu-1125 and Ile-1129 of αC. There is another <scene name='Hepatocyte_growth_factor_receptor/A_and_c_intercalating/2'>interaction</scene> between the residues Ile-1053, Leu-1055 and Leu-1058 of αA and Ile- | + | This structure is made up of many α helical structures that move in the transformation from inactive to active kinase. Some of these helices are conserved in many different tyrosine kinases. C-met does show a divergence from other tyrosine kinases (such as IRK and FGFRK) in the helix formed at the N-terminus, before the core kinase domain, in residues <scene name='Hepatocyte_growth_factor_receptor/1060-1069/1'>1060-1069</scene>. <ref>PMID: 14559966</ref> The αA is in contact with αC and so causes αC to be in a slightly different orientation than in FGFRK and IRK. Residues Leu-1062, Val-1066, and Val-1069 of αA <scene name='Hepatocyte_growth_factor_receptor/A_and_c_intercalating/1'>intercalate</scene> with with residues Leu-1125 and Ile-1129 of αC. There is another <scene name='Hepatocyte_growth_factor_receptor/A_and_c_intercalating/2'>interaction</scene> between the residues Ile-1053, Leu-1055 and Leu-1058 of αA and Ile-1118 and Val-1121 of αC. Because of the movement of αC during activation of the kinase, it is an assumption that αA is also part of the kinase activation upon ligand binding. <ref>PMID: 14559966</ref> |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
==Mutation== | ==Mutation== | ||
| Line 26: | Line 19: | ||
<scene name='User:Juliette_Personius/sandbox_1/1234_and_1235_mutations/1'>tyr1234 and tyr1235</scene> are replaced by a phenylalanine and aspartate, respectively. This mutation normally causes the receptor to be constitutively active, and is found in HNSC (Head, Neck squamous cell) carcinoma. Although there is no longer phosphorylation at these sites, it is believed that the negative charge of the aspartate resembles the negative phosphate that would normally cause activation, and therefore keeps the protein in its active form. <ref>PMID: 14559966</ref> There is a third mutation at Tyr-1194 which is substituted for a <scene name='Hepatocyte_growth_factor_receptor/1194/1'>phenylalanine</scene>. This is shown to point into the <scene name='Hepatocyte_growth_factor_receptor/1198/1'>pocket</scene> formed by Lys-1198 and Leu-1195 from αE. <ref>PMID: 14559966</ref> This structure is conserved in the wild type protein, suggesting that the mutation at residue 1149 is not changing the structure at this position. | <scene name='User:Juliette_Personius/sandbox_1/1234_and_1235_mutations/1'>tyr1234 and tyr1235</scene> are replaced by a phenylalanine and aspartate, respectively. This mutation normally causes the receptor to be constitutively active, and is found in HNSC (Head, Neck squamous cell) carcinoma. Although there is no longer phosphorylation at these sites, it is believed that the negative charge of the aspartate resembles the negative phosphate that would normally cause activation, and therefore keeps the protein in its active form. <ref>PMID: 14559966</ref> There is a third mutation at Tyr-1194 which is substituted for a <scene name='Hepatocyte_growth_factor_receptor/1194/1'>phenylalanine</scene>. This is shown to point into the <scene name='Hepatocyte_growth_factor_receptor/1198/1'>pocket</scene> formed by Lys-1198 and Leu-1195 from αE. <ref>PMID: 14559966</ref> This structure is conserved in the wild type protein, suggesting that the mutation at residue 1149 is not changing the structure at this position. | ||
| - | =K-252a= | + | ==K-252a== |
<scene name='Hepatocyte_growth_factor_receptor/K-252a/1'>K-252a</scene> is a staurosporine analog. Staurosporine is an inhibitor of many Ser/Thr Kinases, and has been shown to also inhibit c-Met activation by inhibiting its autophosphorylation. The structures of K-252a and staurosporine are very similar, with the main difference being that K-252a has a furanose instead of a pyranose structure. The binding of K-252a causes the c-Met to adopt an inhibitory conformation of the A-loop, specifically with residues <scene name='Hepatocyte_growth_factor_receptor/Residues1231-1244/1'>1231-1244</scene> . This segment blocks the place where the substrate tyrosine side chain would bind, if the protein were in an active conformation. Residues | <scene name='Hepatocyte_growth_factor_receptor/K-252a/1'>K-252a</scene> is a staurosporine analog. Staurosporine is an inhibitor of many Ser/Thr Kinases, and has been shown to also inhibit c-Met activation by inhibiting its autophosphorylation. The structures of K-252a and staurosporine are very similar, with the main difference being that K-252a has a furanose instead of a pyranose structure. The binding of K-252a causes the c-Met to adopt an inhibitory conformation of the A-loop, specifically with residues <scene name='Hepatocyte_growth_factor_receptor/Residues1231-1244/1'>1231-1244</scene> . This segment blocks the place where the substrate tyrosine side chain would bind, if the protein were in an active conformation. Residues | ||
| - | <scene name='Hepatocyte_growth_factor_receptor/Catalytic_1127/1'>1223-1230</scene> also enhance this inhibitory conformation, as they constrain αC into a conformation that does not allow the catalytic placement of <scene name='Hepatocyte_growth_factor_receptor/1127/1'>Glu-1127</scene> keeping αC in an inactive conformation. In an active kinase, Glu-1127 would form a salt | + | <scene name='Hepatocyte_growth_factor_receptor/Catalytic_1127/1'>1223-1230</scene> also enhance this inhibitory conformation, as they constrain αC into a conformation that does not allow the catalytic placement of <scene name='Hepatocyte_growth_factor_receptor/1127/1'>Glu-1127</scene> keeping αC in an inactive conformation. In an active kinase, Glu-1127 would form a salt bridge with Lys-1110. |
Residues 1229-1230 pass through the triphosphate subsite of bound ATP blocking ATP binding. The K-252a itself binds in the adenosine pocket, therefore also inhibiting the binding of ATP. The binding of K-252a is very favorable (enthalpy change of -17.9 kcal/mol). This is probably due to polar interactions as well as a change in conformation upon binding. <ref>PMID: 14559966</ref> | Residues 1229-1230 pass through the triphosphate subsite of bound ATP blocking ATP binding. The K-252a itself binds in the adenosine pocket, therefore also inhibiting the binding of ATP. The binding of K-252a is very favorable (enthalpy change of -17.9 kcal/mol). This is probably due to polar interactions as well as a change in conformation upon binding. <ref>PMID: 14559966</ref> | ||
| Line 41: | Line 34: | ||
Met-1229, Met-1211 and Met-1160 all make up the <scene name='Hepatocyte_growth_factor_receptor/Platform/1'>platform</scene> for the indolocarbazole plane as they are all within van der waals distance of it. <ref>PMID: 14559966</ref> | Met-1229, Met-1211 and Met-1160 all make up the <scene name='Hepatocyte_growth_factor_receptor/Platform/1'>platform</scene> for the indolocarbazole plane as they are all within van der waals distance of it. <ref>PMID: 14559966</ref> | ||
| + | ==C-Terminal Docking Site== | ||
| - | + | In c-Met, there are two tyrosines located in the C-terminal tail sequence, which, upon phosphorylation, act as the docking sites for many signal transducers. These tyrosines correspond to residues <scene name='Hepatocyte_growth_factor_receptor/Tyrisine_docking_sites/1'>1349 and 1356</scene>. Both of these sites interact with SH2, MBD and PTD domains of signal transducers. The residues <scene name='Hepatocyte_growth_factor_receptor/Extended_conformation/1'>1349-1352</scene> form an extended conformation, which is seen in other phosphopeptides that bind to SH2 domains. Residues | |
| - | + | <scene name='Hepatocyte_growth_factor_receptor/Beta_1_turn/1'>1353-1356</scene> form a type I β turn, which is similar to sequences that bind to Shc-PTB domians. Whether binding to SH2 domains or PTB domains, upon binding, these motifs would move to avoid clashes with the C lobe. The third binding motif is found in residues <scene name='Hepatocyte_growth_factor_receptor/Type_2_beta_turn/1'>1356-1359</scene>, which form a type II β turn, and is similar to pohsphopeptides that bind Grb2. When comparing the unphosphorylated conformation of the motif to one that is phosphorylated, and bound to the Grb2 complex, there is a peptide flip between the bind of <scene name='Hepatocyte_growth_factor_receptor/1257_and_1258/1'>Val-1357 and Asn-1358</scene>. This suggests that when Grb2 docks onto c-Met, there is a change in orientation of this motif. These three binding motifs of the mutated structure are very similar to binding motifs that would be recognized by their binding partners, implying that the C-terminal supersite of this structure is very similar to that of an active c-met. <ref>PMID: 14559966</ref> | |
| - | In c-Met, there are two tyrosines located in the C-terminal tail sequence, which, upon phosphorylation, act as the docking sites for many signal transducers. These tyrosines correspond to residues <scene name='Hepatocyte_growth_factor_receptor/Tyrisine_docking_sites/1'>1349 and 1356</scene>. Both of these sites interact with SH2, MBD and PTD domains of signal transducers. The residues <scene name='Hepatocyte_growth_factor_receptor/Extended_conformation/1'>1349-1352</scene> form an extended conformation, which is seen in other phosphopeptides that bind to SH2 domains. Residues 1353-1356 form a type I β turn, which is similar to sequences that bind to Shc-PTB domians. Whether binding to SH2 domains or PTB domains, upon binding, these motifs would move to avoid clashes with the C lobe. The third binding motif is found in residues 1356-1359, which form a type II β turn, and is similar to pohsphopeptides that bind Grb2. When comparing the unphosphorylated conformation of the motif to one that is phosphorylated, and bound to the Grb2 complex, there is | + | |
| - | + | ||
==Biological Significance== | ==Biological Significance== | ||
| Line 54: | Line 46: | ||
This structure also shows the binding motifs of c-Met in an unphosphorylated form, giving insight on how the motifs may move when interacting with their respective binding domains (Grb2, SH2, PTB domains). | This structure also shows the binding motifs of c-Met in an unphosphorylated form, giving insight on how the motifs may move when interacting with their respective binding domains (Grb2, SH2, PTB domains). | ||
| + | |||
| + | == 3D Structures of hepatocyte growth factor receptor == | ||
| + | [[Hepatocyte growth factor receptor 3D structures]] | ||
| + | |||
| + | </StructureSection> | ||
==References== | ==References== | ||
<references /> | <references /> | ||
| + | [[Category:Topic Page]] | ||
Current revision
| |||||||||||
References
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ http://en.wikipedia.org/wiki/C-Met
- ↑ Maina F, Casagranda F, Audero E, Simeone A, Comoglio PM, Klein R, Ponzetto C. Uncoupling of Grb2 from the Met receptor in vivo reveals complex roles in muscle development. Cell. 1996 Nov 1;87(3):531-42. PMID:8898205
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
