Monooxygenase
From Proteopedia
(Difference between revisions)
| (12 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
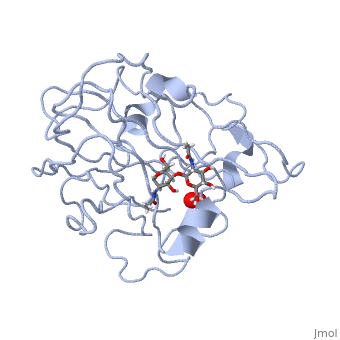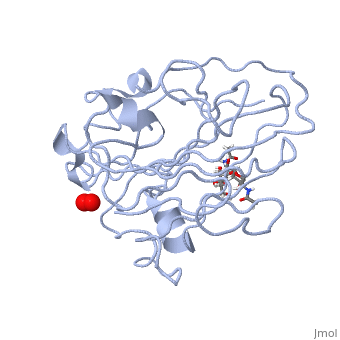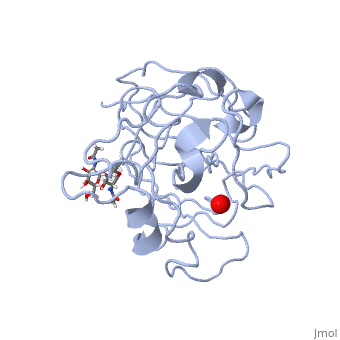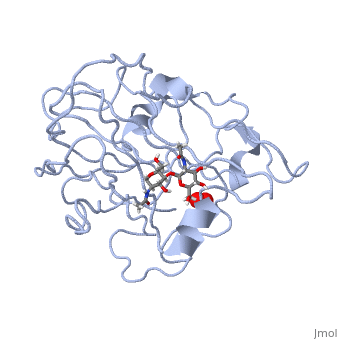
| - | <StructureSection load='5tkh' size='350' side='right' scene='' caption=' | + | <StructureSection load='5tkh' size='350' side='right' scene='' caption='Glycosylated lytic polysaccharide monooxygenase complex with Cu(II) (orange) and peroxide (PDB code [[5tkh]])' > |
| - | + | __TOC__ | |
== Function == | == Function == | ||
'''Monooxygenases''' (MO) catalyzes the incorporation of a hydroxyl group into a variety of substrates. MO catalyzes the reduction of O<sub>2</sub> to H<sub>2</sub>O while oxidating NADPH. | '''Monooxygenases''' (MO) catalyzes the incorporation of a hydroxyl group into a variety of substrates. MO catalyzes the reduction of O<sub>2</sub> to H<sub>2</sub>O while oxidating NADPH. | ||
| + | *'''ActVA-Orf6 monooxygenase''' catalyses the oxidation of an aromatic intermediate of the actinorhodin pathway <ref>PMID:12514126</ref>. | ||
| + | *'''Baeyer-Villigen monooxygenase''' is a bioanalytic tool which can catalyze reactions which are difficult to do via chemical means<ref>PMID:15599520</ref>. | ||
| + | *'''TropB monooxygenase''' catalyses asymmetric oxidative dearomatization reactions <ref>PMID:31346489</ref>. | ||
| + | *'''TetX monooxygenase''' inactivates the tetracycline antibiotic <ref>PMID:15452119</ref>. | ||
| + | *'''Phenol 2-monooxygenase''' see [[Phenol hydroxylase (hebrew)]]. | ||
=== Peptidylglycine α-Hydroxylating Monooxygenase (PHM)-coordination of peroxide to Cu<sub>M</sub> center. Structural and computational study <ref >doi 10.1007/s00775-012-0967-z</ref>=== | === Peptidylglycine α-Hydroxylating Monooxygenase (PHM)-coordination of peroxide to Cu<sub>M</sub> center. Structural and computational study <ref >doi 10.1007/s00775-012-0967-z</ref>=== | ||
| Line 9: | Line 14: | ||
In recent years there has been a significant interest in describing the interactions of copper-containing enzymes with O<sub>2</sub>/H<sub>2</sub>O<sub>2</sub>-derived species. The short-lived intermediates resulting from the activation of dioxygen are the key players in the mechanistic cycles in many metalloenzymes. In the enzyme <scene name='Journal:JBIC:17/Cv/3'>peptidylglycine alpha-hydroxylating monooxygenase (PHM)</scene> various reduced Cu/oxygen species have been proposed to act as catalytically competent intermediates, yet their exact nature and their role in the enzymatic reaction is still unknown. | In recent years there has been a significant interest in describing the interactions of copper-containing enzymes with O<sub>2</sub>/H<sub>2</sub>O<sub>2</sub>-derived species. The short-lived intermediates resulting from the activation of dioxygen are the key players in the mechanistic cycles in many metalloenzymes. In the enzyme <scene name='Journal:JBIC:17/Cv/3'>peptidylglycine alpha-hydroxylating monooxygenase (PHM)</scene> various reduced Cu/oxygen species have been proposed to act as catalytically competent intermediates, yet their exact nature and their role in the enzymatic reaction is still unknown. | ||
Structural and other studies showed that peptidylglycine α-hydroxylating monooxygenase (PHM) contains <scene name='Journal:JBIC:17/Cv/4'>two non-equivalent copper sites (CuH and CuM)</scene>. CuM serves as an oxygen binding and hydrogen abstraction site, CuH is involved in electron transfer. In the structure of Cu(II)-PHM complexed with hydrogen peroxide determined to 1.98 Å resolution, <scene name='Journal:JBIC:17/Cv/7'>(hydro)peroxide binds exclusively to CuM in a slightly asymmetric side-on mode</scene>. The <scene name='Journal:JBIC:17/Cv/8'>interatomic O-O distance of the copper-bound ligand is 1.5, characteristic of peroxide/hydroperoxide species, and the copper-oxygen distances are 2.0 and 2.1</scene> Å. This Cu(II)-bound <scene name='Journal:JBIC:17/Cv/9'>peroxo moiety interacts closely with a molecule of water</scene>, forming <scene name='Journal:JBIC:17/Cv/10'>hydrogen bonds that stabilize the structure</scene>. DFT and QM/MM calculations indicate that this species is a Cu-bound doubly deprotonated peroxidate and that its energy is similar to that of its isomer Cu(I)-bound superoxide. | Structural and other studies showed that peptidylglycine α-hydroxylating monooxygenase (PHM) contains <scene name='Journal:JBIC:17/Cv/4'>two non-equivalent copper sites (CuH and CuM)</scene>. CuM serves as an oxygen binding and hydrogen abstraction site, CuH is involved in electron transfer. In the structure of Cu(II)-PHM complexed with hydrogen peroxide determined to 1.98 Å resolution, <scene name='Journal:JBIC:17/Cv/7'>(hydro)peroxide binds exclusively to CuM in a slightly asymmetric side-on mode</scene>. The <scene name='Journal:JBIC:17/Cv/8'>interatomic O-O distance of the copper-bound ligand is 1.5, characteristic of peroxide/hydroperoxide species, and the copper-oxygen distances are 2.0 and 2.1</scene> Å. This Cu(II)-bound <scene name='Journal:JBIC:17/Cv/9'>peroxo moiety interacts closely with a molecule of water</scene>, forming <scene name='Journal:JBIC:17/Cv/10'>hydrogen bonds that stabilize the structure</scene>. DFT and QM/MM calculations indicate that this species is a Cu-bound doubly deprotonated peroxidate and that its energy is similar to that of its isomer Cu(I)-bound superoxide. | ||
| - | </StructureSection> | ||
==3D structures of monooxygenase== | ==3D structures of monooxygenase== | ||
| + | [[Monooxygenase 3D structures]] | ||
| - | + | </StructureSection> | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
== References == | == References == | ||
Current revision
| |||||||||||
References
- ↑ Sciara G, Kendrew SG, Miele AE, Marsh NG, Federici L, Malatesta F, Schimperna G, Savino C, Vallone B. The structure of ActVA-Orf6, a novel type of monooxygenase involved in actinorhodin biosynthesis. EMBO J. 2003 Jan 15;22(2):205-15. PMID:12514126 doi:http://dx.doi.org/10.1093/emboj/cdg031
- ↑ Fraaije MW, Wu J, Heuts DP, van Hellemond EW, Spelberg JH, Janssen DB. Discovery of a thermostable Baeyer-Villiger monooxygenase by genome mining. Appl Microbiol Biotechnol. 2005 Jan;66(4):393-400. PMID:15599520 doi:10.1007/s00253-004-1749-5
- ↑ Benitez AR, Tweedy S, Baker Dockrey SA, Lukowski AL, Wymore T, Khare D, Brooks CL 3rd, Palfey BA, Smith JL, Narayan ARH. Structural basis for selectivity in flavin-dependent monooxygenase-catalyzed oxidative dearomatization. ACS Catal. 2019 Apr 5;9(4):3633-3640. doi: 10.1021/acscatal.8b04575. Epub 2019, Mar 25. PMID:31346489 doi:http://dx.doi.org/10.1021/acscatal.8b04575
- ↑ Yang W, Moore IF, Koteva KP, Bareich DC, Hughes DW, Wright GD. TetX is a flavin-dependent monooxygenase conferring resistance to tetracycline antibiotics. J Biol Chem. 2004 Dec 10;279(50):52346-52. doi: 10.1074/jbc.M409573200. Epub 2004, Sep 27. PMID:15452119 doi:http://dx.doi.org/10.1074/jbc.M409573200
- ↑ Rudzka K, Moreno DM, Eipper B, Mains R, Estrin DA, Amzel LM. Coordination of peroxide to the Cu(M) center of peptidylglycine alpha-hydroxylating monooxygenase (PHM): structural and computational study. J Biol Inorg Chem. 2012 Dec 18. PMID:23247335 doi:10.1007/s00775-012-0967-z
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, Joel L. Sussman, Alexander Berchansky, Jaime Prilusky
