This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
C-JUN
From Proteopedia
(Difference between revisions)
| (28 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
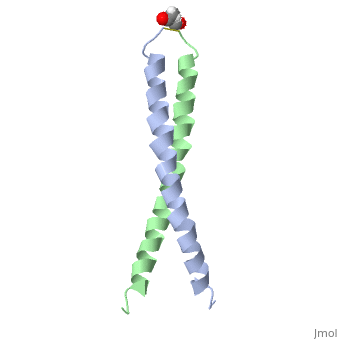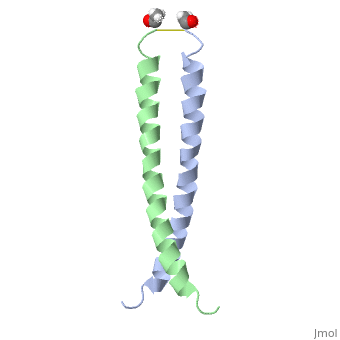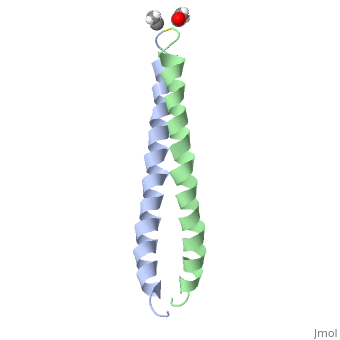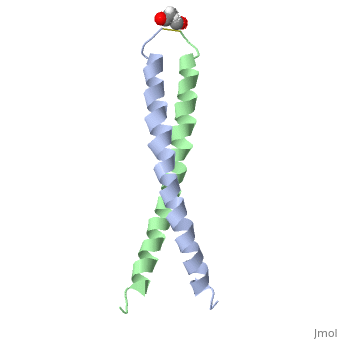
| - | + | <StructureSection load='1jun' size='350' side='right' scene='' caption='Human C-Jun homodimer leucine zipper domain complex with acetyl (PDB code [[1jun]])'> | |
| - | = '''C-JUN''' = | ||
| - | |||
| - | |||
| - | ---- | ||
| - | |||
| - | |||
| - | <applet load='1Z82' size='200' frame='true' align='right' caption='1JUN' /> | ||
== Introduction == | == Introduction == | ||
| - | The c-Jun protein is a member of transcription factors which consist of a basic region leucine zipper region <ref name="one">PMID:8662824</ref>. Originally identified by its homology to v-jun, the oncogene from the avian sarcomoa virus <ref name="four"> Bossy-Wetzel, E., Bakiri, L., Yaniv, M. (1997). Induction of apoptosis by the transcription factor c-Jun. EMO Journal. Vol.16;7. 1695-1709 </ref>. All these leucine zipper factors bind to DNA in one of two states: homo or heterodimers <ref name="two">PMID:8662824</ref>. In conjunction with the c-Fos protein these two proteins bind to specific regions of DNA strands. Together these two proteins form the c-fos/c-jun complex which help regulate cell growth and differentiation <ref name="one">. The members of the jun and fos families include three Jun proteins and four Fos proteins (c-Jun, JunB, JunD,c-Fos, Fos-B, Fra1, and Fra2) <ref name="one">. Regulation of the complex iteslf is done by interactions between the protein and DNA in addition to the protein-protein interactions between each of the leucine zipper domains <ref name="one">. | + | The '''c-Jun''' protein is a member of transcription factors which consist of a basic region leucine zipper region <ref name="one"> PMID:8662824 </ref>. Originally identified by its homology to v-jun, the oncogene from the avian sarcomoa virus <ref name="four"> Bossy-Wetzel, E., Bakiri, L., Yaniv, M. (1997). Induction of apoptosis by the transcription factor c-Jun. EMO Journal. Vol.16;7. 1695-1709 </ref>. All these leucine zipper factors bind to DNA in one of two states: homo or heterodimers <ref name="two"> PMID:8662824 </ref>. In conjunction with the c-Fos protein these two proteins bind to specific regions of DNA strands. Together these two proteins form the c-fos/c-jun complex which help regulate cell growth and differentiation <ref name="one" />. The members of the jun and fos families include three Jun proteins and four Fos proteins (c-Jun, JunB, JunD,c-Fos, Fos-B, Fra1, and Fra2) <ref name="one" />. Regulation of the complex iteslf is done by interactions between the protein and DNA in addition to the protein-protein interactions between each of the leucine zipper domains <ref name="one" />. See [[Transcription and RNA Processing]]. |
== Structure Overview == | == Structure Overview == | ||
| - | + | The structure of c-Jun is comprised of a leucine zipper as previously stated. This dimerization motif may be in one of two classes, both of which are required for DNA-binding transcription factors; the basic-domain leucine zipper proteins (bZIP) and the basic helix loop-helix-leucine zipper proteins(bHLH-ZIP) <ref name="two"> A Junius, F.K., Mackay, J.P., Bubb, W.A., Jensen, S.A., Weiss, A.S., King, G.F. 2006. Nuclear Magnetic Resonance Characterization of the Jun Leucine Zipper Domain: Unusual Properties of Coiled-Coil Interfacial Polar Residues?</ref>. The strand becomes an elongated coiled coil. This is formed by residues at the a and d positions in each of the two monomers, whereby they create hydrophobic centers which conform to the "knobs into holes" model by Crick. <ref name="two" />. Amino acids at these a and d positions are each surrounded by 4 additional residues from adjacent a-helix monomer <ref name="two" />. | |
| - | + | The a and d residues each exhibit varying types of packing in terms of this "knobs into holes" theory. According to Harbury et al.(24) the leucines at the a positions are packed "parallel" in such a way that the C-alpha-C-beta bond vector lies in a parallel manner to the C-alpha-C-alpha vector at the base of the acceptor hole on adjacent helix <ref name="one" />. Whereas the opposite is true for the leucines in the d positions. Here the residues are packed in a "perpendicular" nature <ref name="one" />. The bond vector of the C-alpha-C-beta pack approximately perpendicular to the C-alpha-C-alpha vector at the base of the hole of the second helix in which it packs <ref name="one" />. Therefore only the leucine side chains in the a positions, which point away from the boundary, make van der Waals interactions <ref name="one" />. | |
| - | + | ||
| - | The a and d residues each exhibit varying types of packing in terms of this "knobs into holes" theory. According to Harbury et al.(24) the leucines at the a positions are packed "parallel" in such a way that the C-alpha-C-beta bond vector lies in a parallel manner to the C-alpha-C-alpha vector at the base of the acceptor hole on adjacent helix <ref name="one">. Whereas the opposite is true for the leucines in the d positions. Here the residues are packed in a "perpendicular" nature <ref name="one">. The bond vector of the C-alpha-C-beta pack approximately perpendicular to the C-alpha-C-alpha vector at the base of the hole of the second helix in which it packs <ref name="one">. Therefore only the leucine side chains in the a positions, which point away from the boundary, make van der Waals interactions <ref name="one">. | + | |
== Protein Function == | == Protein Function == | ||
| - | The primary function of c-Jun is in regards to DNA transcription. Specifically, the protein is involved in proliferation, apoptosis, oncogenic transformation and various cellular processes <ref name="three">PMID:12798298</ref>. For instance cells which lack an allele for c-jun have been shown to stunt growth both in vitro and in vivo <ref name="four"/>. Whereas a prolonged and therefore strong induction of c-jun has been in response to such things as tumor necrosis factor or stress inducing stimuli such as ultra violet radiation <ref name="four"/>. | + | The primary function of c-Jun is in regards to DNA transcription. Specifically, the protein is involved in proliferation, apoptosis, oncogenic transformation and various cellular processes <ref name="three"> PMID:12798298 </ref>. For instance cells which lack an allele for c-jun have been shown to stunt growth both in vitro and in vivo <ref name="four" />. Whereas a prolonged and therefore strong induction of c-jun has been in response to such things as tumor necrosis factor or stress inducing stimuli such as ultra violet radiation <ref name="four" />. |
== Protein Regulation == | == Protein Regulation == | ||
| - | Changes made in the phosphorylation state of specific amino acids is one means by which c-Jun regulates transcription <ref name="six"> PMID:8165146 </ref>. To date two seperate sites of phosphorylation have been identified. One is located at the N-terminal end in which the amino acids Ser63 and Ser73 are phosphorylated in response to ''ras'' expression. When ''ras'' is expressed, and Ser63 and Ser73 are phosphorylated,and transcriptional activity of c-Jun increases. The second site is located at the C-terminal which is very close in proximity to the DNA binding domain. Here the residues are Thr214, Ser226, and Ser 232 <ref name="six"/>. Unlike the two serines at the N-terminal end, phosphorylation at the C-terminal end inhibits DNA binding to c-Jun <ref name="six"/>. Therefore with the expression of such oncogenes as ''ras'' dephsphorylation of these three residues occurs. | + | Changes made in the phosphorylation state of specific amino acids is one means by which c-Jun regulates transcription <ref name="six"> PMID:8165146 </ref>. To date two seperate sites of phosphorylation have been identified. One is located at the N-terminal end in which the amino acids Ser63 and Ser73 are phosphorylated in response to ''ras'' expression. When ''ras'' is expressed, and Ser63 and Ser73 are phosphorylated,and transcriptional activity of c-Jun increases. The second site is located at the C-terminal which is very close in proximity to the DNA binding domain. Here the residues are Thr214, Ser226, and Ser 232 <ref name="six" />. Unlike the two serines at the N-terminal end, phosphorylation at the C-terminal end inhibits DNA binding to c-Jun <ref name="six" />. Therefore with the expression of such oncogenes as ''ras'' dephsphorylation of these three residues occurs. |
== Psychological Influences == | == Psychological Influences == | ||
The stress-induced signaling cascade may also active c-Jun by phosphorylation. The N-ternminal protein kinase phosphorylates Ser63 and Ser73 <ref name="five"> PMID:10064599 </ref> . Another mechanism for the activation however is interestingly through intracellular calcium concentrations. Increasing these concentrations by opening the L-type voltage gated calcium channels leads to serines phosphorlation. | The stress-induced signaling cascade may also active c-Jun by phosphorylation. The N-ternminal protein kinase phosphorylates Ser63 and Ser73 <ref name="five"> PMID:10064599 </ref> . Another mechanism for the activation however is interestingly through intracellular calcium concentrations. Increasing these concentrations by opening the L-type voltage gated calcium channels leads to serines phosphorlation. | ||
| - | It was found that the N-terminus contains both calcium and stress-regulated transcriptional activation domains <ref name="five"/>. According to the study,distinct mechanisms of c-Jun control function by calcium and stress signals <ref name="five"/>. | + | It was found that the N-terminus contains both calcium and stress-regulated transcriptional activation domains <ref name="five" />. According to the study,distinct mechanisms of c-Jun control function by calcium and stress signals <ref name="five" />. |
| + | ==Additional Resources== | ||
| + | To See Additional information, see: [[Transcription and RNA Processing]] <br /> | ||
| + | </StructureSection> | ||
| + | ==3D structure of C-JUN== | ||
| + | |||
| + | Updated on {{REVISIONDAY2}}-{{MONTHNAME|{{REVISIONMONTH}}}}-{{REVISIONYEAR}} | ||
| + | |||
| + | [[1jun]] – hCJUN leucine zipper domain – human – NMR<br /> | ||
| + | [[1jnm]] - hCJUN leucine zipper domain + DNA<br /> | ||
| + | [[1fos]] – hCJUN + p55 c-Fos + DNA<br /> | ||
| + | [[5fv8]] – hCJUN + FOSW<br /> | ||
== References == | == References == | ||
<references/> | <references/> | ||
| + | |||
| + | [[Category:Topic Page]] | ||
Current revision
| |||||||||||
3D structure of C-JUN
Updated on 11-April-2018
1jun – hCJUN leucine zipper domain – human – NMR
1jnm - hCJUN leucine zipper domain + DNA
1fos – hCJUN + p55 c-Fos + DNA
5fv8 – hCJUN + FOSW
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 Junius FK, O'Donoghue SI, Nilges M, Weiss AS, King GF. High resolution NMR solution structure of the leucine zipper domain of the c-Jun homodimer. J Biol Chem. 1996 Jun 7;271(23):13663-7. PMID:8662824
- ↑ 2.0 2.1 2.2 Bossy-Wetzel, E., Bakiri, L., Yaniv, M. (1997). Induction of apoptosis by the transcription factor c-Jun. EMO Journal. Vol.16;7. 1695-1709
- ↑ 3.0 3.1 3.2 3.3 Junius FK, O'Donoghue SI, Nilges M, Weiss AS, King GF. High resolution NMR solution structure of the leucine zipper domain of the c-Jun homodimer. J Biol Chem. 1996 Jun 7;271(23):13663-7. PMID:8662824
- ↑ Mechta-Grigoriou F, Giudicelli F, Pujades C, Charnay P, Yaniv M. c-jun regulation and function in the developing hindbrain. Dev Biol. 2003 Jun 15;258(2):419-31. PMID:12798298
- ↑ 5.0 5.1 5.2 Hoeffler WK, Levinson AD, Bauer EA. Activation of c-Jun transcription factor by substitution of a charged residue in its N-terminal domain. Nucleic Acids Res. 1994 Apr 11;22(7):1305-12. PMID:8165146
- ↑ 6.0 6.1 6.2 Cruzalegui FH, Hardingham GE, Bading H. c-Jun functions as a calcium-regulated transcriptional activator in the absence of JNK/SAPK1 activation. EMBO J. 1999 Mar 1;18(5):1335-44. PMID:10064599 doi:10.1093/emboj/18.5.1335
Proteopedia Page Contributors and Editors (what is this?)
Andrew Rebeyka, Michal Harel, Alexander Berchansky, David Canner, Andrea Gorrell
