This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
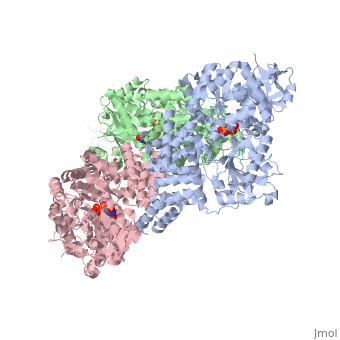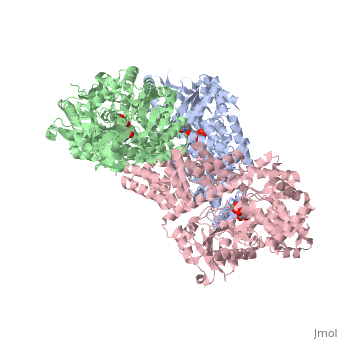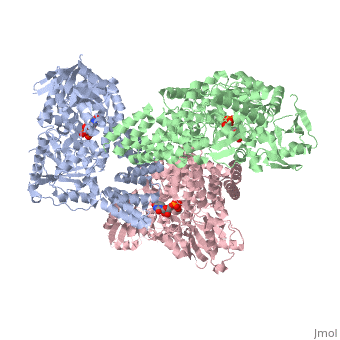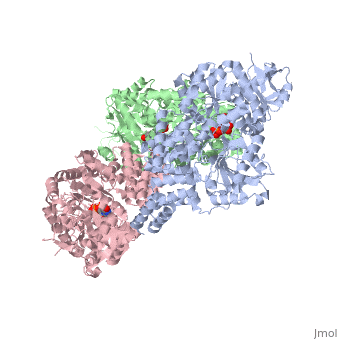
Human O-GlcNAc transferase
From Proteopedia
(Difference between revisions)
(New page: <Structure load='3pe3' size='400' frame='true' align='right' caption='O-GlcNAc transferase with UDP' scene='Insert optional scene name here' /> == O-GlcNAc transferase == O-linked β-...) |
|||
| (7 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
| - | + | <StructureSection load='3pe3' size='450' side='right' scene='' caption=''> | |
| - | < | + | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
O-linked β-N-acetylglucosamine transferase (O-GlcNAc transferase) is an essential mammalian enzyme that acts as a nutrient sensor, coupling metabolic status to the regulation of a wide variety of cellular signaling pathways.<ref> Hart GW, Housley MP, Slawson C. Cycling of O-linked β-N-acetylglucosamine on nucleocytoplasmic proteins. Nature.2007;446:1017-22.[http://www.nature.com/nature/journal/v446/n7139/abs/nature05815.html]</ref> OGT catalyses the transfer of N-acetylglucosamine from UDP-N-acetylglucosamine (UDP-GlcNAc) to serines and threonines of cytoplasmic, nuclear and mitochondrial proteins, including numerous transcription factors, tumour suppressors, kinases, phospahateses and histone-modifying proteins.<ref>PMID:21240259</ref> Two crystal structures of human OGT are reported here as a <scene name='Sandbox_Reserved_381/Ternary_complex_ogt_with_udp/1'>ternary complex</scene> with UDP and a <scene name='Sandbox_Reserved_381/Binary_complex_with_udp/1'>binary complex</scene> with UDP and a peptide substrate. | O-linked β-N-acetylglucosamine transferase (O-GlcNAc transferase) is an essential mammalian enzyme that acts as a nutrient sensor, coupling metabolic status to the regulation of a wide variety of cellular signaling pathways.<ref> Hart GW, Housley MP, Slawson C. Cycling of O-linked β-N-acetylglucosamine on nucleocytoplasmic proteins. Nature.2007;446:1017-22.[http://www.nature.com/nature/journal/v446/n7139/abs/nature05815.html]</ref> OGT catalyses the transfer of N-acetylglucosamine from UDP-N-acetylglucosamine (UDP-GlcNAc) to serines and threonines of cytoplasmic, nuclear and mitochondrial proteins, including numerous transcription factors, tumour suppressors, kinases, phospahateses and histone-modifying proteins.<ref>PMID:21240259</ref> Two crystal structures of human OGT are reported here as a <scene name='Sandbox_Reserved_381/Ternary_complex_ogt_with_udp/1'>ternary complex</scene> with UDP and a <scene name='Sandbox_Reserved_381/Binary_complex_with_udp/1'>binary complex</scene> with UDP and a peptide substrate. | ||
| Line 15: | Line 10: | ||
== OGT Features of Interest == | == OGT Features of Interest == | ||
| - | The OGT protein possess two ligands <scene name='Sandbox_Reserved_381/So4_ligand/1'>SO4</scene> and uridine-5-diphosphate <scene name='Sandbox_Reserved_381/Udp/1'>(UDP)</scene>. OGT is the only known member to glycosylate polypeptides, and it contains a long uncharacterized intervening sequence (~120 amino acids) in the middle of the catalytic region. Studies suggest that OGT contains a phosphatidylinositol (3,4,5)-trisphosphate (PIP<sub>3</sub>)binding domain. The most unusual feature of OGT is the intervening domain between the catalytic lobes, which is only found in metazoans. This polypeptide adopts a topologically novel fold with a seven-stranded <scene name='Sandbox_Reserved_381/Ogt_structure/1'>beta sheet</scene> core stabilized by flanking alpha helices. There are two long <scene name='Sandbox_Reserved_381/Unstructured_loops/1'>unstructured loops</scene> for which electron density is missing.<ref> PMID:18288188</ref> | + | The OGT protein possess two ligands <scene name='Sandbox_Reserved_381/So4_ligand/1'>SO4</scene> and uridine-5-diphosphate <scene name='Sandbox_Reserved_381/Udp/1'>(UDP)</scene>. OGT is the only known member of the GT-B superfamily of glycosyltransferases to glycosylate polypeptides, and it contains a long uncharacterized intervening sequence (~120 amino acids) in the middle of the catalytic region. Studies suggest that OGT contains a phosphatidylinositol (3,4,5)-trisphosphate (PIP<sub>3</sub>)binding domain. The most unusual feature of OGT is the intervening domain between the catalytic lobes, which is only found in metazoans. This polypeptide adopts a topologically novel fold with a seven-stranded <scene name='Sandbox_Reserved_381/Ogt_structure/1'>beta sheet</scene> core stabilized by flanking alpha helices. There are two long <scene name='Sandbox_Reserved_381/Unstructured_loops/1'>unstructured loops</scene> for which electron density is missing.<ref> PMID:18288188</ref> |
== OGT Structure == | == OGT Structure == | ||
| Line 21: | Line 16: | ||
== N Terminus == | == N Terminus == | ||
| - | < | + | <scene name='47/475977/Cv/1'>Superhelical TPR domain of OGT, structural similarities to importin alpha</scene> (PDB entry [[1w3b]]). |
| + | |||
| + | The crystal structure of the homodimeric TPR domain of human OGT, which contains 11.5 TPR repeats gives insight into the mechanism of target recognition. The repeats form an elongated superhilix. The concave surface of the superhelix is lined by absolutely conserved asparagines, in a manner reminiscent of the peptide-binding site of importin alpha. Based on this structural similarity, it is proposed that OGT uses an analogous molecular mechanism to recognize its targets.<ref>PMID:15361863</ref> | ||
== OGT Mediated Disease == | == OGT Mediated Disease == | ||
Faulty regulation of O-GlcNAc modifications has been suggested to be involved in neurodegenerative diseases, diabetes mellitus and cancer. Biochemical details of these processes are still unclear.<ref>PMID:16051707</ref> Proteins modified by O-GlcNAc have been directly shown to have a role in the pathology of human diseases. For instance, the Ser/The kinase AKT,PI<sub>(3)</sub>K,insulin receptor substrate 1, glycogen synthase and endothelial nitric oxide synthase,all of which are enzymes that have a crucial role in insulin signalling,are reversibly modified by OGT. A recent study showed that recruitment of OGT to the plasma membrane specifically prevents the phosphorylation of AKT and possibly other proteins, thereby terminating insulin signalling.<ref>PMID:18288188</ref> This adds evidence to the view that increasing the level of O-GlcNAc modifications correlates with the development of insulin resistance, which is a characteristic of type II diabetes.<ref>PMID:16317114</ref><ref>PMID:16781888</ref> Some indications suggest that O-GlcNAc modifications have a role in Alzheimer disease. Higher levels of O-GlcNAc can be detected in the brain tissue, and several proteins involved in neuronal signaling are modified with O-GlcNAc. Among them are the β-amyloid precursor protein, clathrin-assembly proteins and neurofilaments. In the brains of patients with Alzheimer disease, hyperphosphorylated Tau protein was modified by O-GlcNAc to a lesser extent than in healthy individuals.<ref>PMID:17940659</ref> Studies have shown that some oncogenes and tumour suppressors are targets of O-glycosylation, including the SV40 T antigen and c-MYC.<ref>PMID:14533811</ref> Tumour cells have an altered glucose metabolism that is expected to produce changes in O-GlcNAc levels and to affect different signaling pathways. | Faulty regulation of O-GlcNAc modifications has been suggested to be involved in neurodegenerative diseases, diabetes mellitus and cancer. Biochemical details of these processes are still unclear.<ref>PMID:16051707</ref> Proteins modified by O-GlcNAc have been directly shown to have a role in the pathology of human diseases. For instance, the Ser/The kinase AKT,PI<sub>(3)</sub>K,insulin receptor substrate 1, glycogen synthase and endothelial nitric oxide synthase,all of which are enzymes that have a crucial role in insulin signalling,are reversibly modified by OGT. A recent study showed that recruitment of OGT to the plasma membrane specifically prevents the phosphorylation of AKT and possibly other proteins, thereby terminating insulin signalling.<ref>PMID:18288188</ref> This adds evidence to the view that increasing the level of O-GlcNAc modifications correlates with the development of insulin resistance, which is a characteristic of type II diabetes.<ref>PMID:16317114</ref><ref>PMID:16781888</ref> Some indications suggest that O-GlcNAc modifications have a role in Alzheimer disease. Higher levels of O-GlcNAc can be detected in the brain tissue, and several proteins involved in neuronal signaling are modified with O-GlcNAc. Among them are the β-amyloid precursor protein, clathrin-assembly proteins and neurofilaments. In the brains of patients with Alzheimer disease, hyperphosphorylated Tau protein was modified by O-GlcNAc to a lesser extent than in healthy individuals.<ref>PMID:17940659</ref> Studies have shown that some oncogenes and tumour suppressors are targets of O-glycosylation, including the SV40 T antigen and c-MYC.<ref>PMID:14533811</ref> Tumour cells have an altered glucose metabolism that is expected to produce changes in O-GlcNAc levels and to affect different signaling pathways. | ||
| + | </StructureSection> | ||
| + | __NOTOC__ | ||
| + | |||
| + | ==3D structures of O-GlcNAc transferase== | ||
| + | |||
| + | [[O-GlcNAc transferase]] | ||
==References== | ==References== | ||
<references /> | <references /> | ||
Current revision
| |||||||||||
3D structures of O-GlcNAc transferase
References
- ↑ Hart GW, Housley MP, Slawson C. Cycling of O-linked β-N-acetylglucosamine on nucleocytoplasmic proteins. Nature.2007;446:1017-22.[1]
- ↑ Lazarus MB, Nam Y, Jiang J, Sliz P, Walker S. Structure of human O-GlcNAc transferase and its complex with a peptide substrate. Nature. 2011 Jan 27;469(7331):564-7. Epub 2011 Jan 16. PMID:21240259 doi:10.1038/nature09638
- ↑ Hart GW, Housley MP, Slawson C. Cycling of O-linked beta-N-acetylglucosamine on nucleocytoplasmic proteins. Nature. 2007 Apr 26;446(7139):1017-22. PMID:17460662 doi:10.1038/nature05815
- ↑ Yang X, Ongusaha PP, Miles PD, Havstad JC, Zhang F, So WV, Kudlow JE, Michell RH, Olefsky JM, Field SJ, Evans RM. Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature. 2008 Feb 21;451(7181):964-9. PMID:18288188 doi:10.1038/nature06668
- ↑ Cavill I. Reducing blood transfusion. Focus should be on improving patients' ability to make own blood. BMJ. 2002 Sep 21;325(7365):655. PMID:12269319
- ↑ Gambetta MC, Oktaba K, Muller J. Essential role of the glycosyltransferase sxc/Ogt in polycomb repression. Science. 2009 Jul 3;325(5936):93-6. Epub 2009 May 28. PMID:19478141 doi:10.1126/science.1169727
- ↑ Alexander G, Danilo G. The O-linked N-acetylglucosamine modification in cellular signalling and the immune system. EMBO reports. 2008 June;9:748-753[2]
- ↑ Yang X, Ongusaha PP, Miles PD, Havstad JC, Zhang F, So WV, Kudlow JE, Michell RH, Olefsky JM, Field SJ, Evans RM. Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature. 2008 Feb 21;451(7181):964-9. PMID:18288188 doi:10.1038/nature06668
- ↑ Kreppel LK, Blomberg MA, Hart GW. Dynamic glycosylation of nuclear and cytosolic proteins. Cloning and characterization of a unique O-GlcNAc transferase with multiple tetratricopeptide repeats. J Biol Chem. 1997 Apr 4;272(14):9308-15. PMID:9083067
- ↑ Kreppel L, Hart G. Regulation of a cytosolic and nuclear O-GlcNAc transferase. Role of the tetratricopeptide repeats. J Biol Chem. 1999;274:32015-32022
- ↑ Jinek M, Rehwinkel J, Lazarus BD, Izaurralde E, Hanover JA, Conti E. The superhelical TPR-repeat domain of O-linked GlcNAc transferase exhibits structural similarities to importin alpha. Nat Struct Mol Biol. 2004 Oct;11(10):1001-7. Epub 2004 Sep 12. PMID:15361863 doi:10.1038/nsmb833
- ↑ Hanover JA, Forsythe ME, Hennessey PT, Brodigan TM, Love DC, Ashwell G, Krause M. A Caenorhabditis elegans model of insulin resistance: altered macronutrient storage and dauer formation in an OGT-1 knockout. Proc Natl Acad Sci U S A. 2005 Aug 9;102(32):11266-71. Epub 2005 Jul 28. PMID:16051707 doi:10.1073/pnas.0408771102
- ↑ Yang X, Ongusaha PP, Miles PD, Havstad JC, Zhang F, So WV, Kudlow JE, Michell RH, Olefsky JM, Field SJ, Evans RM. Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature. 2008 Feb 21;451(7181):964-9. PMID:18288188 doi:10.1038/nature06668
- ↑ Love DC, Hanover JA. The hexosamine signaling pathway: deciphering the "O-GlcNAc code". Sci STKE. 2005 Nov 29;2005(312):re13. PMID:16317114 doi:10.1126/stke.3122005re13
- ↑ Zachara NE, Hart GW. Cell signaling, the essential role of O-GlcNAc! Biochim Biophys Acta. 2006 May-Jun;1761(5-6):599-617. Epub 2006 May 6. PMID:16781888 doi:10.1016/j.bbalip.2006.04.007
- ↑ Dias WB, Hart GW. O-GlcNAc modification in diabetes and Alzheimer's disease. Mol Biosyst. 2007 Nov;3(11):766-72. Epub 2007 Aug 29. PMID:17940659 doi:10.1039/b704905f
- ↑ Chou TY, Hart GW. O-linked N-acetylglucosamine and cancer: messages from the glycosylation of c-Myc. Adv Exp Med Biol. 2001;491:413-8. PMID:14533811
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, David Canner, Sheri Marcum, Alexander Berchansky
