|
|
| (463 intermediate revisions not shown.) |
| Line 3: |
Line 3: |
| | <!-- INSERT YOUR SCENES AND TEXT BELOW THIS LINE --> | | <!-- INSERT YOUR SCENES AND TEXT BELOW THIS LINE --> |
| | | | |
| - | =='''EcoRV endonuclease'''== | + | =='''Vitamin D receptor/vitamin D (1db1)<ref>PMID: 10678179 </ref>'''== |
| | + | by Roger Crocker, Kate Daborowski, Patrick Murphy, Benjamin Rizkin and Aaron Thole |
| | | | |
| - | <Structure load='1rva' size='500' frame='true' align='right' caption='EcoRV is a restriction enzyme found in escherichia coli bacteria that cleaves DNA and helps to protect the cell against foreign DNA' scene='Insert optional scene name here' /> | |
| | | | |
| - | ===Introduction=== | + | [[Student Projects for UMass Chemistry 423 Spring 2016]] |
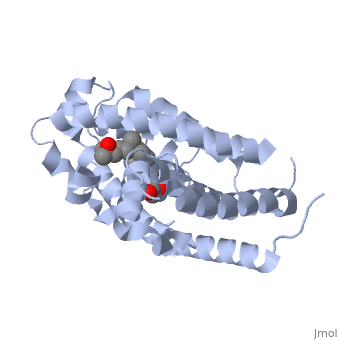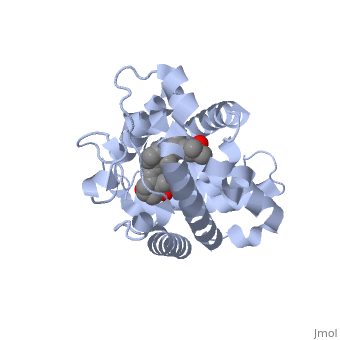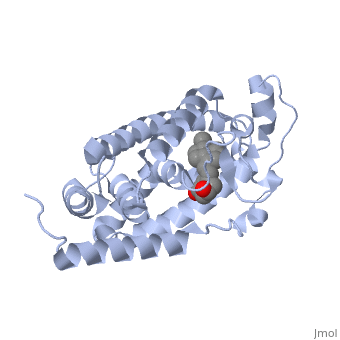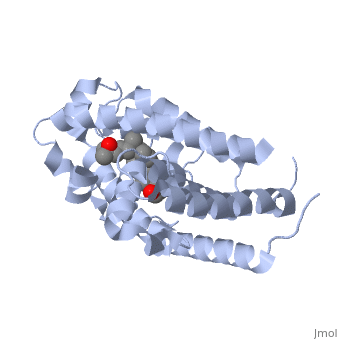
| | + | <StructureSection load='1db1' size='350' side='right' caption='caption for Molecular Playground (PDB entry [[1db1]])' scene=''> |
| | | | |
| - | EcoRV endonuclease is a type II restriction enzyme, or restriction endonuclease, found in e. coli bacteria. The main biological function of restriction endonucleases is to protect the cell's genome against any foreign DNA. Restriction enzymes recognize and cleave specific sequences of DNA. In the figure to the right, the enzyme is shown with the <scene name='Sandbox_Reserved_428/Dna_sequence/1'>eleven base sequence</scene> AAAGATATCTT. A type II restriction enzymes cleave at a short distance from the recognition site and often use Mg(2+) as a cofactor, as does this enzyme. They are commonly found in bacteria and shared structural features indicate that they are evolutionarily related. Type II endonucleases have been the site of much research because of applications in gene analysis and cloning and because they are great at modeling protein-DNA interactions.
| + | ==Introduction== |
| | + | <br> |
| | + | The <scene name='48/483885/Color1/6'>vitamin D receptor</scene> (VDR) is a ligand-dependent transcriptional regulator with two strands. VDR belongs to the superfamily of nuclear receptors which control homeostasis, cell differentiation and growth, and many physiological processes. All proteins that belong to the nuclear receptor superfamily have a variable N-terminus region (A/B region), a hinge region that is flexible (D region), a conserved DNA-binding region (DBD, C region), and a moderately conserved ligand-binding region (LBD, E/F region). In the case of VDR, the A/B region is very short so it does not have any AF-1 function and the ligand binding region has a dimerization interface and a transcriptional activation domain that is ligand-dependent (AF-2).[1] <br> <br> |
| | + | The VDR has both an active and suppressed form. The activation or suppression function is caused by the binding of the DR3 response element as a heterodimer with the retinoid X receptor of the target genes. Due to the interactions with the basal transcriptional machinery and transcriptional cofactors, transcription is either activated or suppressed. When VDR is in its active form it regulates both phosphate and calcium metabolism, has immunosuppressive effects, and induces cell differentiation. When there are defects in the VDR that effect its metabolism it can lead to diseases such as severe rickets, secondary hyperparathyroidism, and hypocalcemia. Though defects in VDR can cause many diseases, fully functioning VDR can be used as treatment for disease such as cancer, autoimmune disease, psoriasis, osteoporosis, and renal osteodystrophy.[1] |
| | | | |
| - | Specifically, EcoRV is an orthodox restriction endonuclease. This means that the DNA sequence recognized is palindromic, meaning each strand contains the same sequence. The DNA duplex is cleaved at the phosphodiester bond located at 5'-GAT*ATC-3'. The other DNA strand will also be cleaved in the same location, producing blunt ends. This is relatively unique among restriction enzymes, as many cleave each DNA stand at a different location, leaving what are known as sticky ends. Cleavage occurs by the breaking of the bond between a 3' oxygen and the phosphorus by nucleophilic attack by water. The acidic residues Asp74 and Asp90 are located near the phosphodiester group and provide ligands for the activating Mg(2+) cofactor. | |
| | | | |
| - | The steps of DNA cleavage are as follows. EcoRV binds to the DNA without specificity, which is followed by a diffusional walking "search" down the DNA molecule. If the protein encounters its recognition site, conformational changes occur in the enzyme-DNA complex. The binding to a <scene name='Sandbox_Reserved_428/Non_specific/1'>Non specific</scene> section of DNA is looser, and when the protein encounters the <scene name='Sandbox_Reserved_428/Specific/1'>correct sequence</scene> of DNA it tightens and bends the DNA strands approximately 50 degrees. The DNA is cleaved and released from the enzyme. | |
| | | | |
| | + | ==Overall Structure== |
| | + | The <scene name='48/483885/Alphabeta_picture/1'>vitamin d receptor</scene> contains 427 amino acids with a total molecular weight of 48,289 Da. The protein is also composed almost entirely of alpha helices with only a single beta sheet. The vitamin D receptor also does not have a quaternary structure [2]. <scene name='48/483885/Vitamin_d3/2'>Vitamin D3</scene> is a large organic compound made up of 27 carbon atoms, 44 hydrogen atoms and a single oxygen atom, with the ligand having a total molecular weight of 385 Da [3]. In studying the vitamin d receptor, the regions of the protein have been categorized into domains, with the A/B domain located at the N-terminus, the C domain, which is located between amino acid 20 and amino acid 115, the D domain, which is located between the end of the C domain and amino acid 220, and the EF domain, which encompasses the rest of the protein [4]. |
| | + | <br> <br> |
| | + | Other aspects of interest about the vitamin D receptor include the protein revealing a binding pocket when it is in its active folded state, allowing the ligand to bind to the receptor. The ligand interacts with the activation helix by stabilizing the agonist position. This is accomplished through Van der Waals interactions between the ligand and the activation helix. The activation ligand is a nuclear receptor. There is also some empty space observed around the aliphatic chain [1]. |
| | + | The protein has an active conformation of 1,25 (OH)_2D_3 that has a ligand binding pocket in its active folded state. The activation ligand, a nuclear receptor (VDR), interacts with with the activation helix by stabilizing the agonist position. This is accomplished through Van der Waals interactions between the ligand and the activation helix. There is some empty space observed around the aliphatic chain, indicating the presence of water to stabilize all possible hydrogen bonds [1]. |
| | | | |
| | + | ==Binding Interactions== |
| | + | Protein 1db1 is found to complex with 1,25 Dihydroxy <scene name='48/483885/Vitamin_d3/1'>vitamin D3</scene>. This molecule has three notable alcohol groups shown in red. Oxygen is electronegative, giving alcohols the ability to participate in hydrogen bonding with the protein. Vitamin D3 has a large number of relations with the residues on the protein chain. First in the sequence are <scene name='48/483885/Residues_140-151/2'>residues 140-151</scene>. Tyr143, shown in blue, is the closest to the ligand at 2.83 angstroms. This is sightly large but there is still the possibility of hydrogen bonding. Tyr147 in green and Phe150 in black are also known to have interactions with vitamin D3 they are farther away and therefore less significant. Next down the peptide chain are <scene name='48/483885/Residues_235-240/1'>residues 235-240</scene>. Ser237, shown in green, has significant interactions with vitamin D3 this can be seen by its short distance 2.78 angstroms. Only 40 residues away, more hydrogen bonding is occurring.<scene name='48/483885/Residues_270-280/1'>Arg274 and Ser278</scene> form bonds with the same oxygen atoms as Tyr143 and Ser237 respectively. This means that the oxygen atoms of vitamin D3, when bound to the receptor, are negatively charged and stabilized by protons. On the opposite end of vitamin D3 there is an additional negatively charged oxygen. Although this oxygen does not participate in hydrogen bonding. <scene name='48/483885/His305_and_his397/1'>His305 and His397</scene>, shown in blue and green respectively, Contain aromatic rings. These rings are able to momentarily accept the electrons donated by the oxygen because they can delocalize the charge. This creates two pseudo-covalent bonds that is approximately 2.81 angstroms. When looking at <scene name='48/483885/All_binding_interactions/1'>all binding interactions</scene>, it can be seen that all of the binding sights are centered around oxygen [5,6]. |
| | | | |
| | + | ==Additional Features== |
| | | | |
| | + | Hereditary Vitamin D Resistant Rickets (HVDRR) is a condition that most commonly occurs in children and results in soft and weak bone formation, which often causes deformities in bone structure. A lack of proper nutrients, Vitamin D3 in particular, as well as defects in the Vitamin D receptor can cause rickets in humans. HVDRR can occur when the VDR is impaired in its ability to activate transcription in response to the 1,25-(OH)2D3 ligand [7]. VDR regulates the hormonal form of Vitamin D, through modifying the transcription of the target to a certain sequence of DNA, called the Vitamin D responsive element (VDRE). This activation requires an additional receptor, which is a Retinoid X Receptor (RXR) to bind to the heterodimer [7]. A mutation in the transcription of the protein has the potential to result in this disease and the mutation results in the <scene name='48/483885/Heterodimer/1'>heterodimer</scene> not forming properly. Current research shows that there are two mechanisms that can cause this failed transcription and inability to bind to the heterodimer to occur within the body. The first is a point mutation of an amino acid in the zinc finger region of the VDR that reduces the binding of the heterodimer, this region is found in the amino acid residues 21-85 [7]. The other is a premature stop codon in the DNA sequences that does not allow for the full transcription, which can have an effect of reducing the affinity of the heterodimer binding [7]. Although there may be other mutations that could cause HVDRR to occur within the body, these two types of mutations have been shown to be major causes of the disease. |
| | | | |
| - | <scene name='Sandbox_Reserved_428/Non_specific/1'>Non specific DNA</scene>
| + | Also, the VDR has an effect on the hair follicle cycle, which has been observed through the eliminating the expression of this receptor. In null-VDR mice, it has been shown that with normal mineral ion levels that the mice result in alopecia, a disease that causes hair loss [8]. VDR is expressed in the hair follicle keratinocytes and its levels are higher in the late anagen and catagen stages of the hair cycle [8]. These two stages are vital in the differentiation and proliferation of hair follicle keratinocytes, which regulate hair growth in the body. Much research has been done into the mechanism in which the VDR effects the hair follicle cycle with the overall mechanism still unknown. The mechanism was first believed to be that of binding the VDR to 1,25- dihydroxyvitamin D causing transactivation due to the fact that targeted expressions of wild-type VDR to the keratinocytes of VDR null mice rescued alopecia [8]. Although, this was disproven through investigations in vitamin D3-deficient mice that had no detectable 1,25-dihydroxyvitamin D for the VDR to bind, yet the mice did not develop alopecia. This shows that the VDR transcriptional activation of DNA is not the main cause of the loss of hair follicles. Current research observes the ligand-independent actions of the VDR that have not been observed extensively as a mechanism [8]. Nuclear receptor co-repressor genes have been observed in studies to have an effect on the hair follicle cycle including the HR gene (Hairless). This corepressor has been shown to have interactions with the VDR in vivo and tests with the mutation of Hairless have caused alopecia in mice in vivo [8]. Thus, although the mechanism behind the interaction of Hairless and the VDR is still unknown it has been shown in studies that there is a relationship between the two in the body and that it has been linked to the hair follicle cycle. |
| - | <scene name='Sandbox_Reserved_428/Specific/1'>Specific</scene>
| + | |
| - | <scene name='Sandbox_Reserved_428/Active_sites/1'>Active sites</scene>
| + | |
| - | <scene name='Sandbox_Reserved_428/Dna_sequence/1'>TextToBeDisplayed</scene>
| + | |
| - | <scene name='Sandbox_Reserved_428/Dna_zoom/1'>DNA zoom</scene>
| + | |
| | | | |
| - | (I'll delete these before the presentation, its just easier to work with them all in one spot) | |
| | | | |
| - | <font color='red'>Very interesting! A few suggestions:
| + | ==Quiz Question 1== |
| - | Tell us and use green scenes to show us which part of the DNA sequence is the recognition site and where is the cleavage site. Use color in the text to make it easier to quickly see your points: for example color the text and the DNA molecule red for the recognition sequence, then use color in both the text and structure to highlight the cleavage site.
| + | Osteoporosis is a disease in which the bones become porous and fragile. The most common cause of the ailment is calcium deficiency. As the vitamin D receptor has association with calcium uptake, mutations in VDR could be detrimental.If an individual had a point mutation that would replace <scene name='48/483885/Point_mutation/1'>His305 and His397</scene> with serine amino groups, would that individual be more likely to develop osteoporosis? why? |
| | + | ==See Also== |
| | + | *[[3w0a]] |
| | + | *[[3w0c]] |
| | + | *[[3w0y]] |
| | + | *[[3w5t]] |
| | + | *[[3w5r]] |
| | + | *[[1db1]] |
| | | | |
| - | If Mg2+ is present show it to us (space fill) and color "Mg" in the text the same color as in the structure.
| + | ==Credits== |
| | | | |
| - | If Mg is not present, is this to keep the enzyme from cleaving the DNA? That would be an interesting point to comment on.
| + | Introduction - Kate Daborowski |
| | | | |
| - | You could try coloring the DNA and text to make the palindrome clear.
| + | Overall Structure - Aaron Thole and Benjamin Rizkin |
| | | | |
| - | If you want to describe the active site, use colored text and a green scene to show us your points (or you could leave this to another section, or help with that section).
| + | Drug Binding Site - Roger Crocker |
| | | | |
| - | What are the structures shown in your non-specific and correct sequence green scenes? Are these different pdb file? Please include the pdb codes. You make very interesting points about the binding, but need better green scenes to explain this change (e.g. show the DNA bend and other conformational changes). I suggest passing these off to the Additional Features section, which currently has no green scenes and could make improved versions of these and fully explain the change.
| + | Additional Features - Patrick Murphy |
| | | | |
| - | I bet you could make a cooler caption for the molecular playground, about scissors for cutting DNA and cloning... but keep it similar length or shorter.
| + | Quiz Question 1 - Roger Crocker |
| | + | ==References== |
| | + | <references/> |
| | | | |
| - | Length of your section is fine (about the length of jmol window): do not increase.
| + | 1. Rochel N, Wurtz JM, Mitschler A, Klaholz B, Moras D. The crystal structure of the nuclear receptor for vitamin D bound to its natural ligand. Mol Cell. 2000 Jan;5(1):173-9. |
| - | </font>
| + | |
| | | | |
| - | ===Overall Structure===
| + | 2. VDR Gene http://www.genecards.org/cgi-bin/carddisp.pl?gene=VDR (accessed Apr 2, 2016). |
| - | <Structure load='1rva' size='500' frame='true' align='right' caption='EcoRV' scene='Insert optional scene name here' />
| + | |
| - | EcoRV endonuclease is functional as a dimer consisting of two monomers; both monomers depicted <scene name='Sandbox_Reserved_428/U-shaped_dimer/1'>here</scene> are shown in green or purple in a U shape. Each monomer, monomer A shown <scene name='Sandbox_Reserved_428/Monomera/1'>here</scene> and monomer B shown <scene name='Sandbox_Reserved_428/Monomer/1'>here</scene>, consists of 244 amino acids arranged in alpha/beta secondary structures, but the monomers are not identical.
| + | |
| | | | |
| - | Monomer B, previously shown in green, has 9 alpha helices, shown <scene name='Sandbox_Reserved_428/Alpha_helix_-_monomer/1'>here</scene>, in pink and 11 beta strands, shown <scene name='Sandbox_Reserved_428/Beta_strands_-_monomer/1'>here</scene>, in blue. The beta strands form 3 beta sheets of various sizes, both parallel and anti parallel. The <scene name='Sandbox_Reserved_428/Largest_beta_sheet_-_monomer/1'>largest beta sheet</scene> is a mixture of parallel and anti-parallel strands. The light purple and dark purple strands make up two anti-parallel sheets. The connection between the dark and light purple beta sheets is parallel. Depicted <scene name='Sandbox_Reserved_428/Tripleap_beta_sheets_-_monomer/1'>here</scene> is a short triple-stranded antiparallel sheet that helps form the top side of each monomer. Opposite this beta sheet,shown <scene name='Sandbox_Reserved_428/Apattachbeta_strands_-_monomer/4'>here</scene>, on the bottom of the enzyme, is another triple strand anti-parallel beta sheet.
| + | 3. Vitamin D3 https://pubchem.ncbi.nlm.nih.gov/compound/Vitamin_D3#section=2D-Structure (accessed Apr 10, 2016). |
| | | | |
| - | Monomer A, previously shown in purple has 10 alpha helices, shown <scene name='Sandbox_Reserved_428/Alpha_helix_-_monomera/1'>here</scene>, in pink and 10 beta stands, shown <scene name='Sandbox_Reserved_428/Beta_sheets_-_monomera/1'>here</scene>, in blue. The differences between monomer A and monomer B lies only in the connection between the two monomers. At the point of connection a 5 strand beta sheet, made of 3 stands from monomer B and 2 strands from monomer A, this beta sheet assists in the structure and stability of the dimer as a whole. Shown <scene name='Sandbox_Reserved_428/Beta_sheets_-_dimer/1'>here</scene>, the green beta sheet is from monomer A and the purple beta sheet is from monomer B. Once you <scene name='Sandbox_Reserved_428/Zoom_out_beta_sheets_-_dimer/1'>zoom out</scene>, you can see how these two anti-parallel beta sheets form one anti-parallel beta sheet connecting the two monomers at the bottom of the U shape. Monomer A also has an extra alpha helix, residue number 144-150, shown <scene name='Sandbox_Reserved_428/Diff_btwn_a_and_b/1'>here</scene>. For visual reference the monomer A and B connecting beta sheet is shown in dark purple (A beta strands) and dark green (B beta strands).
| + | 4. Strugnell, S.; Deluca, H. The Vitamin D Receptor - Structure and Transcriptional Activation. Experimental Biology and Medicine1997, 215, 223–228. |
| | | | |
| - | There are two structural sub-domains. The first, called the <scene name='Sandbox_Reserved_428/Dimerization_sub-domain/1'>dimerization sub-domain</scene>, is rather small and forms most of the dimer interface, residue numbers 19-32 and 150-160. The second is called the <scene name='Sandbox_Reserved_428/Dna-binding_subdomain/1'>DNA binding sub-domain</scene>, residue numbers 2-18 38-140 and 167-243. The remaining segments of amino acids make up a <scene name='Sandbox_Reserved_428/Sub-domain_linkage/1'>flexible linkage</scene> the two sub-domains, residue numbers 33-37,141-149, and 161-165.
| + | 5. Vitamin D Receptor http://pdb101.rcsb.org/motm/155 (accessed Apr 4, 2016). |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | ===Binding Interactions===
| + | |
| - | <Structure load='1rva' size='500' frame='true' align='right' caption='Insert caption here' scene='Insert optional scene name here' />
| + | |
| - | <scene name='Sandbox_Reserved_428/Binds_to_dna/1'>DNA Binding</scene>
| + | |
| - | | + | |
| - | DNA recognition sites on the EcoRV molecule, called R-loops, bind to the major grooves of the double stranded DNA at its recognition sequence GATATC by hydrogen bonding. This enzyme is a type II restriction endonuclease, which means this enzyme cleaves the DNA recognition sequence at the center (between the T and A base pairs). These hydrogen bonds makes the DNA form a kinked conformation that is later stabilized by the addition of the Mg2+ ion. The Mg2+ ion is a catalyst that causes the DNA to shift in a way that increases the rate necessary for DNA cleavage.
| + | |
| - | | + | |
| - | The Mg2+ binding site is formed when ionic interactions cause the slightly acidic Asp90 residue and the slightly negatively charged scissile phosphodiester group to approach each other. This allows the Mg2+ ion to bind to this enzyme, also with ionic interactions between the positively charged Mg2+ and the partially negative charged oxygen atoms from multiple molecules. These molecules that bind to the Mg2+ ion are the carboxylate oxygen atoms from the Asp74 and Asp90 residues, the nonesterified oxygen from the scissile phosphodiester group, and three additional oxygen atoms from three water molecules. These six ionic bonds form an octahedral shape in the active site of this enzyme.
| + | |
| - | | + | |
| - | These six ionic interactions all have about the same binding distance except for one bond between the oxygen from the Asp74 residue and the Mg2+ ion that is significantly longer. The five similar bond lengths are all about 2.08 Å, but the bond between Mg2+ and the Asp74 oxygen spans a distance of 2.9 Å. This is noted because the Asp90 and scissile phosphodiester molecules that bind to this Mg2+ ion change their bonding interactions with hydrogen to accommodate the addition of the Mg2+ ion. The Asp74 residue maintains its hydrogen bond interactions on its side chain with the main chain of the Ile91 residue and the water molecule, which is why it keeps a greater distance between itself and the Mg2+ ion.
| + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | ===Additional Features===
| + | |
| - | <Structure load='1rva' size='500' frame='true' align='right' caption='Insert caption here' scene='Insert optional scene name here' />
| + | |
| - | | + | |
| - | EcoRV does not require ATP hydrolysis for its nucleolytic activity. The type II isozyme can be further classified according to its recognition site, presence of the divalent Magnesium cofactor, and the interactions present about the active site. EcoRV DNA cleavage is an excellent example of protein induced DNA conformational change: DNA bends > unstacking of bases > widening of minor groove > compression of the major groove > Scissile phosphates are brought deeper into the active site. furthermore, bending is conserved following cleavage. The precise mechanism of cleavage is yet to be established. Participation of magnesium is of research interest due to the activity of the enzyme occurring only with the presence of magnesium or calcium.
| + | |
| - | | + | |
| - | The enzyme Specificity for the respective DNA sequence is a result of conformational changes of both the enzyme and DNA, both of which occur as the enzyme approaches the recognition sequence and allows for more intimate contact with cleavage site. Presence of magnesium relieves repulsive electrostatic interactions between the protein and DNA, this is not necessary in other restriction enzymes due to the presence of carboxylates within the active sites groups which can be protonated.
| + | |
| - | | + | |
| - | ===Credits===
| + | |
| - | | + | |
| - | Introduction - Jesse
| + | |
| - | | + | |
| - | Overall Structure - Nicole
| + | |
| - | | + | |
| - | Drug Binding Site - Julia
| + | |
| - | | + | |
| - | Additional Features - Sam
| + | |
| - | | + | |
| - | ===References===
| + | |
| - | <references/>
| + | |
| - | 1. Kostrewa D, Winkler FK. Mg2+ binding to the active site of EcoRV endonuclease: a crystallographic study of complexes with substrate and product DNA at 2 A resolution. Biochemistry. 1995 Jan 17;34(2):683-96. PMID:7819264
| + | |
| | | | |
| - | 2. Berg, J. ''Biochemistry,'' 7th edition.
| + | 6. 3D Binding Pocket http://www.rcsb.org/pdb/explore/jmol.do?structureId=1DB1&residueNr=VDX (accessed Apr 4, 2016). |
| | | | |
| - | 3. Winkler, Fritz K. The crystal structure of EcoRV endonuclease and of its complexes with cognate and non-cognate DNA fragments. The EMBO Journal 12, p1781-1795. 1993
| + | 7. Whitfield, G.K.; Selznick, S.H.; Haussler, C.A.; Hsieh, J.; Galligan, M.A.; Jurutka, P.W.; Thompson, P.D.; Lee, S.M.; Zerwekh, J.E.; Haussler, M.R. Vitamin D Receptors from Patients with Resistance to 1,25-Dihydroxyvitamin D3: Point Mutations Confer Reduced Transactivation in Response to Ligand and Impair Interaction with the Retinoid X Receptor Heterodimeric Partner. Mol Endocrinol, 1996 10 (12): 1617-1631 |
| | | | |
| - | 4. Pingoud A., Jeltsch A. Structure and function of type II restriction endonucleases. ''Nucleic Acids Research''. 29 (18)3705–3727.
| + | 8. Skorija, K.; Cox, M.; Sisk, J.M.; Dowd, D.R.; MacDonald, P.N.; Thompson, C.C.; Demay, M.B. Ligand- Independent Actions of the Vitamin D Receptor Maintain Hair Follicle Homeostasis. Mol Endocrinol, April 2005, 19(4):855–862 |
by Roger Crocker, Kate Daborowski, Patrick Murphy, Benjamin Rizkin and Aaron Thole
|
Introduction
The (VDR) is a ligand-dependent transcriptional regulator with two strands. VDR belongs to the superfamily of nuclear receptors which control homeostasis, cell differentiation and growth, and many physiological processes. All proteins that belong to the nuclear receptor superfamily have a variable N-terminus region (A/B region), a hinge region that is flexible (D region), a conserved DNA-binding region (DBD, C region), and a moderately conserved ligand-binding region (LBD, E/F region). In the case of VDR, the A/B region is very short so it does not have any AF-1 function and the ligand binding region has a dimerization interface and a transcriptional activation domain that is ligand-dependent (AF-2).[1]
The VDR has both an active and suppressed form. The activation or suppression function is caused by the binding of the DR3 response element as a heterodimer with the retinoid X receptor of the target genes. Due to the interactions with the basal transcriptional machinery and transcriptional cofactors, transcription is either activated or suppressed. When VDR is in its active form it regulates both phosphate and calcium metabolism, has immunosuppressive effects, and induces cell differentiation. When there are defects in the VDR that effect its metabolism it can lead to diseases such as severe rickets, secondary hyperparathyroidism, and hypocalcemia. Though defects in VDR can cause many diseases, fully functioning VDR can be used as treatment for disease such as cancer, autoimmune disease, psoriasis, osteoporosis, and renal osteodystrophy.[1]
Overall Structure
The contains 427 amino acids with a total molecular weight of 48,289 Da. The protein is also composed almost entirely of alpha helices with only a single beta sheet. The vitamin D receptor also does not have a quaternary structure [2]. is a large organic compound made up of 27 carbon atoms, 44 hydrogen atoms and a single oxygen atom, with the ligand having a total molecular weight of 385 Da [3]. In studying the vitamin d receptor, the regions of the protein have been categorized into domains, with the A/B domain located at the N-terminus, the C domain, which is located between amino acid 20 and amino acid 115, the D domain, which is located between the end of the C domain and amino acid 220, and the EF domain, which encompasses the rest of the protein [4].
Other aspects of interest about the vitamin D receptor include the protein revealing a binding pocket when it is in its active folded state, allowing the ligand to bind to the receptor. The ligand interacts with the activation helix by stabilizing the agonist position. This is accomplished through Van der Waals interactions between the ligand and the activation helix. The activation ligand is a nuclear receptor. There is also some empty space observed around the aliphatic chain [1].
The protein has an active conformation of 1,25 (OH)_2D_3 that has a ligand binding pocket in its active folded state. The activation ligand, a nuclear receptor (VDR), interacts with with the activation helix by stabilizing the agonist position. This is accomplished through Van der Waals interactions between the ligand and the activation helix. There is some empty space observed around the aliphatic chain, indicating the presence of water to stabilize all possible hydrogen bonds [1].
Binding Interactions
Protein 1db1 is found to complex with 1,25 Dihydroxy . This molecule has three notable alcohol groups shown in red. Oxygen is electronegative, giving alcohols the ability to participate in hydrogen bonding with the protein. Vitamin D3 has a large number of relations with the residues on the protein chain. First in the sequence are . Tyr143, shown in blue, is the closest to the ligand at 2.83 angstroms. This is sightly large but there is still the possibility of hydrogen bonding. Tyr147 in green and Phe150 in black are also known to have interactions with vitamin D3 they are farther away and therefore less significant. Next down the peptide chain are . Ser237, shown in green, has significant interactions with vitamin D3 this can be seen by its short distance 2.78 angstroms. Only 40 residues away, more hydrogen bonding is occurring. form bonds with the same oxygen atoms as Tyr143 and Ser237 respectively. This means that the oxygen atoms of vitamin D3, when bound to the receptor, are negatively charged and stabilized by protons. On the opposite end of vitamin D3 there is an additional negatively charged oxygen. Although this oxygen does not participate in hydrogen bonding. , shown in blue and green respectively, Contain aromatic rings. These rings are able to momentarily accept the electrons donated by the oxygen because they can delocalize the charge. This creates two pseudo-covalent bonds that is approximately 2.81 angstroms. When looking at , it can be seen that all of the binding sights are centered around oxygen [5,6].
Additional Features
Hereditary Vitamin D Resistant Rickets (HVDRR) is a condition that most commonly occurs in children and results in soft and weak bone formation, which often causes deformities in bone structure. A lack of proper nutrients, Vitamin D3 in particular, as well as defects in the Vitamin D receptor can cause rickets in humans. HVDRR can occur when the VDR is impaired in its ability to activate transcription in response to the 1,25-(OH)2D3 ligand [7]. VDR regulates the hormonal form of Vitamin D, through modifying the transcription of the target to a certain sequence of DNA, called the Vitamin D responsive element (VDRE). This activation requires an additional receptor, which is a Retinoid X Receptor (RXR) to bind to the heterodimer [7]. A mutation in the transcription of the protein has the potential to result in this disease and the mutation results in the not forming properly. Current research shows that there are two mechanisms that can cause this failed transcription and inability to bind to the heterodimer to occur within the body. The first is a point mutation of an amino acid in the zinc finger region of the VDR that reduces the binding of the heterodimer, this region is found in the amino acid residues 21-85 [7]. The other is a premature stop codon in the DNA sequences that does not allow for the full transcription, which can have an effect of reducing the affinity of the heterodimer binding [7]. Although there may be other mutations that could cause HVDRR to occur within the body, these two types of mutations have been shown to be major causes of the disease.
Also, the VDR has an effect on the hair follicle cycle, which has been observed through the eliminating the expression of this receptor. In null-VDR mice, it has been shown that with normal mineral ion levels that the mice result in alopecia, a disease that causes hair loss [8]. VDR is expressed in the hair follicle keratinocytes and its levels are higher in the late anagen and catagen stages of the hair cycle [8]. These two stages are vital in the differentiation and proliferation of hair follicle keratinocytes, which regulate hair growth in the body. Much research has been done into the mechanism in which the VDR effects the hair follicle cycle with the overall mechanism still unknown. The mechanism was first believed to be that of binding the VDR to 1,25- dihydroxyvitamin D causing transactivation due to the fact that targeted expressions of wild-type VDR to the keratinocytes of VDR null mice rescued alopecia [8]. Although, this was disproven through investigations in vitamin D3-deficient mice that had no detectable 1,25-dihydroxyvitamin D for the VDR to bind, yet the mice did not develop alopecia. This shows that the VDR transcriptional activation of DNA is not the main cause of the loss of hair follicles. Current research observes the ligand-independent actions of the VDR that have not been observed extensively as a mechanism [8]. Nuclear receptor co-repressor genes have been observed in studies to have an effect on the hair follicle cycle including the HR gene (Hairless). This corepressor has been shown to have interactions with the VDR in vivo and tests with the mutation of Hairless have caused alopecia in mice in vivo [8]. Thus, although the mechanism behind the interaction of Hairless and the VDR is still unknown it has been shown in studies that there is a relationship between the two in the body and that it has been linked to the hair follicle cycle.
Quiz Question 1
Osteoporosis is a disease in which the bones become porous and fragile. The most common cause of the ailment is calcium deficiency. As the vitamin D receptor has association with calcium uptake, mutations in VDR could be detrimental.If an individual had a point mutation that would replace with serine amino groups, would that individual be more likely to develop osteoporosis? why?
See Also
Credits
Introduction - Kate Daborowski
Overall Structure - Aaron Thole and Benjamin Rizkin
Drug Binding Site - Roger Crocker
Additional Features - Patrick Murphy
Quiz Question 1 - Roger Crocker
References
- ↑ Rochel N, Wurtz JM, Mitschler A, Klaholz B, Moras D. The crystal structure of the nuclear receptor for vitamin D bound to its natural ligand. Mol Cell. 2000 Jan;5(1):173-9. PMID:10678179
1. Rochel N, Wurtz JM, Mitschler A, Klaholz B, Moras D. The crystal structure of the nuclear receptor for vitamin D bound to its natural ligand. Mol Cell. 2000 Jan;5(1):173-9.
2. VDR Gene http://www.genecards.org/cgi-bin/carddisp.pl?gene=VDR (accessed Apr 2, 2016).
3. Vitamin D3 https://pubchem.ncbi.nlm.nih.gov/compound/Vitamin_D3#section=2D-Structure (accessed Apr 10, 2016).
4. Strugnell, S.; Deluca, H. The Vitamin D Receptor - Structure and Transcriptional Activation. Experimental Biology and Medicine1997, 215, 223–228.
5. Vitamin D Receptor http://pdb101.rcsb.org/motm/155 (accessed Apr 4, 2016).
6. 3D Binding Pocket http://www.rcsb.org/pdb/explore/jmol.do?structureId=1DB1&residueNr=VDX (accessed Apr 4, 2016).
7. Whitfield, G.K.; Selznick, S.H.; Haussler, C.A.; Hsieh, J.; Galligan, M.A.; Jurutka, P.W.; Thompson, P.D.; Lee, S.M.; Zerwekh, J.E.; Haussler, M.R. Vitamin D Receptors from Patients with Resistance to 1,25-Dihydroxyvitamin D3: Point Mutations Confer Reduced Transactivation in Response to Ligand and Impair Interaction with the Retinoid X Receptor Heterodimeric Partner. Mol Endocrinol, 1996 10 (12): 1617-1631
8. Skorija, K.; Cox, M.; Sisk, J.M.; Dowd, D.R.; MacDonald, P.N.; Thompson, C.C.; Demay, M.B. Ligand- Independent Actions of the Vitamin D Receptor Maintain Hair Follicle Homeostasis. Mol Endocrinol, April 2005, 19(4):855–862
|