This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
Beta secretase
From Proteopedia
(Difference between revisions)
| (40 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
| - | + | <StructureSection load='' size='350' side='right' scene='Beta_secretase/Default3/1' caption='Human β-secretase (PDB code [[1w51]])'> | |
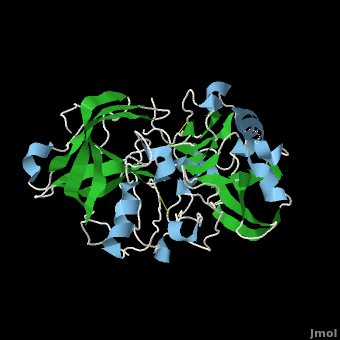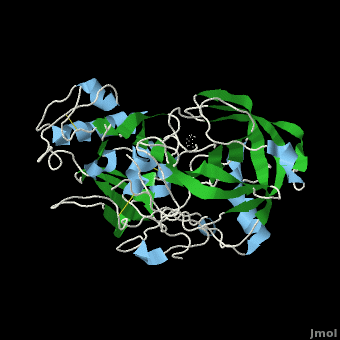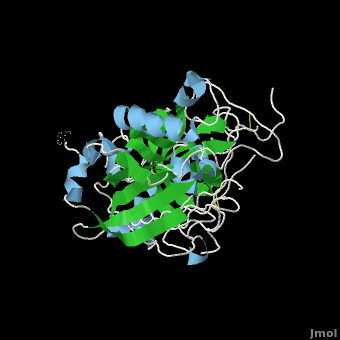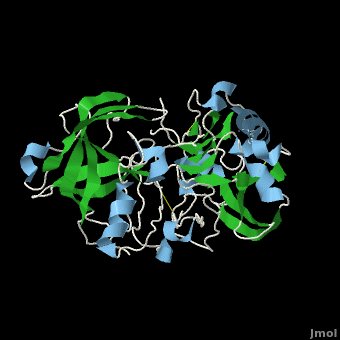
| - | <scene name='Beta_secretase/Default3/1'>β-Secretase</scene>, also known as '''BACE''' ('''β'''-site of '''A'''myloid precursor protein '''C'''leaving '''E'''nzyme) and '''Memapsin 2''', is an aspartyl protease found in humans. β-Secretase plays an important role in the development of [[Alzheimer's Disease]]. This has made β-secretase a therapeutic target for pharmacological intervention. | + | <scene name='Beta_secretase/Default3/1'>β-Secretase</scene>, also known as '''BACE''' ('''β'''-site of '''A'''myloid precursor protein '''C'''leaving '''E'''nzyme) and '''Memapsin 2''', is an aspartyl protease found in humans. β-Secretase plays an important role in the development of [[Alzheimer's Disease]]. This has made β-secretase a therapeutic target for pharmacological intervention. See also<br /> |
| + | *[[Beta-secretase 1]]<br /> | ||
| + | *[[Beta Secretase (BACE1) 1SGZ]]<br /> | ||
| + | *[[Beta Secretase (Arabic)]]<br /> | ||
| + | *[[Beta Secretase (Hebrew)]] . | ||
| + | *'''BACE1''' is expressed mostly in brain and pancreas. | ||
| + | *'''BACE2''' is expressed mostly in kidney tissue. | ||
__TOC__ | __TOC__ | ||
| Line 12: | Line 18: | ||
The other important feature is the <scene name='Beta_secretase/10sloop2/2'>10s loop made up of residues 9 through 14</scene>. The 10s loop is located in the S3 pocket of β-secretase, right between two β strands. When the 10s loop takes an open conformation, it allows for greater binding between the substrate and the S3 pocket. The 10s loop also contains within it a <scene name='Beta_secretase/10sloop2/3'>glycine residue (Gly11)</scene> with which the substrate can form a hydrogen bond, allowing for further stabilization of the 10s loop, as well as the overall β-secretase-substrate interaction. | The other important feature is the <scene name='Beta_secretase/10sloop2/2'>10s loop made up of residues 9 through 14</scene>. The 10s loop is located in the S3 pocket of β-secretase, right between two β strands. When the 10s loop takes an open conformation, it allows for greater binding between the substrate and the S3 pocket. The 10s loop also contains within it a <scene name='Beta_secretase/10sloop2/3'>glycine residue (Gly11)</scene> with which the substrate can form a hydrogen bond, allowing for further stabilization of the 10s loop, as well as the overall β-secretase-substrate interaction. | ||
| - | [[Image:Beta secretase2.png| | + | [[Image:Beta secretase2.png|left|430px]] |
| - | + | {{Clear}} | |
The three parts come together to form a sort of binding pocket for β-secretase's substrates or inhibitors. Binding to the active site activates the flap to close and initiates binding by the 10s loop, all to help stabilize the structure. | The three parts come together to form a sort of binding pocket for β-secretase's substrates or inhibitors. Binding to the active site activates the flap to close and initiates binding by the 10s loop, all to help stabilize the structure. | ||
| Line 21: | Line 27: | ||
Alzheimer's disease occurs by plaques in the walls of vessels and extracellular parenchyma in the brain that lead to the death of neurons. The plaques themselves are formed from the buildup of amyloid beta (Aβ). Aβ is a fragment of about 43 amino acids, and is produced by the cleavage of amyloid precursor protein (APP), an integral membrane protein found in the synapses of neurons. The cleavage of APP itself can go down one of two pathways: | Alzheimer's disease occurs by plaques in the walls of vessels and extracellular parenchyma in the brain that lead to the death of neurons. The plaques themselves are formed from the buildup of amyloid beta (Aβ). Aβ is a fragment of about 43 amino acids, and is produced by the cleavage of amyloid precursor protein (APP), an integral membrane protein found in the synapses of neurons. The cleavage of APP itself can go down one of two pathways: | ||
| - | [[Image:APP2.png| | + | [[Image:APP2.png|left|430px]] |
| - | + | {{Clear}} | |
A. In the first pathway, α-secretase cleaves APP somewhere within the Aβ region. This creates a fragment known as sAPPα. This fragment is beneficial to neurons as it helps to protect them. γ-Secretase can also follow up and cleave at its target, forming a fragment known as p3, whose function is unknown. | A. In the first pathway, α-secretase cleaves APP somewhere within the Aβ region. This creates a fragment known as sAPPα. This fragment is beneficial to neurons as it helps to protect them. γ-Secretase can also follow up and cleave at its target, forming a fragment known as p3, whose function is unknown. | ||
| Line 34: | Line 40: | ||
==Inhibition of Beta Secretase== | ==Inhibition of Beta Secretase== | ||
| - | <applet load="1w51" size="300" color="white" frame="true" align="left" caption="β-secretase complexed with OM99-2" scene="Beta_secretase/Om99-2again/1" /> | ||
| - | |||
Due to β-secretase's function in the production of Aβ, it has become a very popular target for therapeutic drugs. An example of one such developed drug is <scene name='Beta_secretase/Om99-2again/1'>OM99-2</scene> (<scene name='Beta_secretase/Om99-2itself/1'>a hydroxyethylamine compount</scene>), which comes from Astex Technology. | Due to β-secretase's function in the production of Aβ, it has become a very popular target for therapeutic drugs. An example of one such developed drug is <scene name='Beta_secretase/Om99-2again/1'>OM99-2</scene> (<scene name='Beta_secretase/Om99-2itself/1'>a hydroxyethylamine compount</scene>), which comes from Astex Technology. | ||
| - | [[Image:OM99-2.PNG|thumb| | + | [[Image:OM99-2.PNG|thumb|left|'''Fig. 1'''. Structure of OM99-2.]] |
Once the inhibitor moves into place, its positively charged amine group and its hydroxyl group start to interact with β-secretase's active site. The nucelophilic attack on the aspartate's carbonyls binds OM99-2 to β-secretase. As OM99-2 becomes situated within β-secretase's binding pocket, the flap closes upon OM99-2. The flap's residues Thr72 and Gln73 bind with one of OM99-2's carbonyl groups. The 10s loop remains open to allow OM99-2 to interact with the S3 pocket. Gly11 also forms a hydgrogen bond using its carbonyl with the amino terminus of OM99-2. At this point OM99-2 is locked securely within <scene name='Beta_secretase/Om99-2final/1'>β-secretase's binding pocket</scene>. | Once the inhibitor moves into place, its positively charged amine group and its hydroxyl group start to interact with β-secretase's active site. The nucelophilic attack on the aspartate's carbonyls binds OM99-2 to β-secretase. As OM99-2 becomes situated within β-secretase's binding pocket, the flap closes upon OM99-2. The flap's residues Thr72 and Gln73 bind with one of OM99-2's carbonyl groups. The 10s loop remains open to allow OM99-2 to interact with the S3 pocket. Gly11 also forms a hydgrogen bond using its carbonyl with the amino terminus of OM99-2. At this point OM99-2 is locked securely within <scene name='Beta_secretase/Om99-2final/1'>β-secretase's binding pocket</scene>. | ||
| - | [[Image:Om99 reaction.PNG| | + | [[Image:Om99 reaction.PNG|left|430px|thumb]] |
| - | + | {{Clear}} | |
OM99-2 remains stabilized in the pocket by binding to the other pockets of β-secretase (S1, S2, S2', and S3). OM99-2 complexed with β-secretase shows the hydrophobic nature of the S1 and S3 pockets. Additionally, OM99-2 is able to interact with other residues in β-secretase (in this case Gly34 and Thr232), though this is not the case for all inhibitors. | OM99-2 remains stabilized in the pocket by binding to the other pockets of β-secretase (S1, S2, S2', and S3). OM99-2 complexed with β-secretase shows the hydrophobic nature of the S1 and S3 pockets. Additionally, OM99-2 is able to interact with other residues in β-secretase (in this case Gly34 and Thr232), though this is not the case for all inhibitors. | ||
| Line 50: | Line 54: | ||
The apo structure of β-secretase was obtained by refolding recombinant protein developed with ''E. coli''. The β-secretase/OM99-2 crystal structure was obtained by soaking β-secretase with OM99-2 and stabilizing the entire interaction with iodide ions. Mass spectrometry and X-ray crytstallography were used to analyze the obtained structures. | The apo structure of β-secretase was obtained by refolding recombinant protein developed with ''E. coli''. The β-secretase/OM99-2 crystal structure was obtained by soaking β-secretase with OM99-2 and stabilizing the entire interaction with iodide ions. Mass spectrometry and X-ray crytstallography were used to analyze the obtained structures. | ||
| - | <StructureSection load='W50.pdb' size='500' side='right' scene='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/2' caption=''> | ||
== Flexibility of the flap in the active site of BACE1 as revealed by crystal structures and molecular dynamics simulations == | == Flexibility of the flap in the active site of BACE1 as revealed by crystal structures and molecular dynamics simulations == | ||
| - | <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/3'>β-Secretase (β-site amyloid precursor protein cleaving enzyme 1; BACE1)</scene> is a transmembrane aspartic protease that cleaves the β-amyloid precursor protein en route to generation of the amyloid β-peptide (Aβ) that is believed to be the principal component of the amyloid plaques seen in the brains of Alzheimer’s disease. It is thus a prime target for the development of inhibitors that may serve as drugs in the treatment and/or prevention of Alzheimer’s disease. The protease domain has a conserved aspartic protease fold, with the <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/4'>substrate-binding cleft located between the N- and C-terminal lobes</scene> (colored < | + | <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/3'>β-Secretase (β-site amyloid precursor protein cleaving enzyme 1; BACE1)</scene> is a transmembrane aspartic protease that cleaves the β-amyloid precursor protein en route to generation of the amyloid β-peptide (Aβ) that is believed to be the principal component of the amyloid plaques seen in the brains of Alzheimer’s disease. It is thus a prime target for the development of inhibitors that may serve as drugs in the treatment and/or prevention of Alzheimer’s disease. The protease domain has a conserved aspartic protease fold, with the <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/4'>substrate-binding cleft located between the N- and C-terminal lobes</scene> (colored <span style="color:cyan;background-color:black;font-weight:bold;">cyan</span> and <span style="color:yellow;background-color:black;font-weight:bold;">yellow</span>, respectively). <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/5'>The conserved catalytic Asp dyad, Asp32 and Asp228, is located at the interface of the two lobes.</scene> The hairpin loop in the N-terminal lobe that is known as the <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/6'>‘flap’ (residues 67-75) partially covers the cleft, and is perpendicular to it</scene>, as is in general the case for the aspartic protease family. Determination of the crystal structures of both apo and complexed BACE1, structural analysis of all crystal structures of BACE1 deposited in the PDB, and MD simulations of <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/7'>monomeric</scene> and <scene name='Journal:Acta_Crystallogr_D_Biol_Crystallogr:1/Cv/9'>‘dimeric’</scene> BACE1 were used to study conformational changes in the active-site region of the enzyme (the <font color='magenta'><b>first monomer in magenta</b></font> with a <font color='red'><b>red flap</b></font>, the <span style="color:lime;background-color:black;font-weight:bold;">second one in green</span> with an <span style="color:orange;background-color:black;font-weight:bold;">orange flap</span>). Four new crystal structures were determined: that of a <scene name='Journal:Acta_Cryst_D:1/Cv1/1'>BSIIV/BACE1 complex belonging to space group</scene> P6<sub>1</sub>22 ([[3tpr]], <font color='magenta'><b>BACE1 in magenta</b></font> and <scene name='Journal:Acta_Cryst_D:1/Cv1/2'>BSIIV</scene> in <span style="color:cyan;background-color:black;font-weight:bold;">cyan</span>), those of a <scene name='Journal:Acta_Cryst_D:1/Cv1/3'>second BSIIV/BACE1 complex</scene> ([[3tpp]], <font color='indigo'><b>BACE1 in indigo</b></font> and <scene name='Journal:Acta_Cryst_D:1/Cv1/4'>BSIIV</scene> in <span style="color:lime;background-color:black;font-weight:bold;">green</span>) and of an <scene name='Journal:Acta_Cryst_D:1/Cv1/5'>apo structure</scene> ([[3tpj]], in <span style="color:orange;background-color:black;font-weight:bold;">orange</span>), both belonging to space group C222<sub>1</sub>, and that of a <scene name='Journal:Acta_Cryst_D:1/Cv1/6'>second apo structure belonging to space group C2</scene> ([[3tpl]], in <font color='blueviolet'><b>blueviolet</b></font>). <scene name='Journal:Acta_Cryst_D:1/Cv1/1'>The crystals of the BSIIV complex belonging to space group</scene> P6<sub>1</sub>22 ([[3tpr]], <span style="color:cyan;background-color:black;font-weight:bold;">BSIIV</span>/<font color='magenta'><b>BACE1</b></font>) were obtained by <scene name='Journal:Acta_Cryst_D:1/Cv2/6'>soaking the inhibitor BSIIV into a crystal of the apo enzyme belonging to the same space group</scene>, while those of the complex belonging to space group C222<sub>1</sub> were obtained by <scene name='Journal:Acta_Cryst_D:1/Cv2/7'>cocrystallization of the inhibitor and the enzyme</scene> ([[3tpp]], <span style="color:lime;background-color:black;font-weight:bold;">BSIIV</span>/<font color='indigo'><b>BACE1</b></font>). The two apo structures belonging to different space groups were obtained by cocrystallization in the presence of another low affinity inhibitor, using two different precipitants; in both cases the inhibitor was not seen in the crystals thus obtained. It was found that the <scene name='Journal:Acta_Cryst_D:1/Cv2/8'>conformations of the enzyme in the two BSIIV/BACE1 crystal structures differ significantly from each other</scene>, as do the <scene name='Journal:Acta_Cryst_D:1/Cv1/8'>two apo structures</scene>, particularly in the <scene name='Journal:Acta_Cryst_D:1/Cv2/11'>active-site flap and in the 10S and 113S loops</scene>. This led to a systematic analysis of all the BACE1 crystal structures deposited in the PDB (both apo structures and complexes), and to perform MD simulations on the apo enzyme, so as to investigate the underlying causes for the various conformations adopted by the flap. In particular, an attempt was made to clarify whether they might be a consequence of crystal packing in different space groups, or might indicate that the active site can assume multiple low-energy conformations. The RMSFs of main-chain atoms, calculated based on all the BACE1 crystal structures deposited in the PDB, show that the <scene name='Journal:Acta_Cryst_D:1/Cv4/1'>flap is the most flexible region of the protein</scene>. The <scene name='Journal:Acta_Cryst_D:1/Cv5/2'>minimal distance between the flap residues and the catalytic residue D228, which provides a measure of the extent of opening of the flap</scene>, reveals that the <scene name='Journal:Acta_Cryst_D:1/Cv5/3'>flap adopts an open conformation in all the apo structures</scene> (''e.g.'' [[1w50]], <font color='deepskyblue'><b>colored deepskyblue</b></font>) with two exceptions; not surprisingly, the situation with respect to the structures of the complexes is more complex. <scene name='Journal:Acta_Cryst_D:1/Cv5/4'>In most of the complexes, direct H-bond interaction between main chain atoms of flap residues and the bound inhibitor lock the flap in the closed conformation</scene> (''e.g.'' [[3tpp]], <span style="color:lime;background-color:black;font-weight:bold;">BSIIV</span>/<font color='indigo'><b>BACE1</b></font>), except in a few cases in which the position and orientation of the inhibitors sterically preclude its closing. It is of interest that <scene name='Journal:Acta_Cryst_D:1/Cv5/6'>no direct H-bonds are formed between the inhibitor and the flap in complexes in which the flap is in a more open conformation, as is the case in the crystal structure of the complex obtained by soaking BSIIV into apo crystals of BACE1</scene> ([[3tpr]], <span style="color:cyan;background-color:black;font-weight:bold;">BSIIV</span>/<font color='magenta'><b>BACE1</b></font>); but a direct H-bond between Q73 and BSIIV locks the flap in a closed conformation in the co-crystallized structure ([[3tpp]], <span style="color:lime;background-color:black;font-weight:bold;">BSIIV</span>/<font color='indigo'><b>BACE1</b></font>). It may thus be concluded that formation of a direct H-bond between a main-chain atom of a flap residue and the bound inhibitor is a prerequisite for the flap to assume a closed conformation in a complex. <scene name='Journal:Acta_Cryst_D:1/Cv5/2'>So, there are three possible conformations of flap - locked, open and intermediate.</scene> Interestingly, <scene name='Journal:Acta_Cryst_D:1/Cv6/4'>in all the crystal structures of complexes that belong to space group</scene> P6<sub>1</sub>22 except one, the flap is in an open conformation. Our soaked crystals of the BSIIV/BACE1 complex also belong to space group P6<sub>1</sub>22. Analysis of the crystal packing pattern reveals that in this space group the flap is at a packing interface. MD simulations were carried out to check if the observed packing would influence the conformation of the flap. In the starting model for MD that includes a symmetry-related copy of BACE1, both the degree of opening and the time that the flap dwells in the open state are much larger than in the trajectories generated for the monomer. Thus, the packing pattern in crystals belonging to space group P6<sub>1</sub>22 hinders the conformational change in the flap, which may be the main reason why in our <span style="color:cyan;background-color:black;font-weight:bold;">BSIIV</span>/<font color='magenta'><b>BACE1</b></font> complex ([[3tpr]]) belonging to space group P6<sub>1</sub>22 the <scene name='Journal:Acta_Cryst_D:1/Cv5/6'>bound BSIIV does not cause the flap to adopt a closed conformation, whereas it does so in the complex belonging to space group</scene> C222<sub>1</sub> ([[3tpp]], <span style="color:lime;background-color:black;font-weight:bold;">BSIIV</span>/<font color='indigo'><b>BACE1</b></font>). Thus a systematic study of the conformational flexibility of BACE1 may not only contribute to structure-based drug design of BACE1 inhibitors, but also produce a better understanding of the mechanistic events associated with binding of substrates and inhibitors to the enzyme. |
| - | + | ||
| - | + | ||
| - | + | ||
== 3D Structures of Beta secretase == | == 3D Structures of Beta secretase == | ||
| + | [[Beta secretase 3D structures]] | ||
| - | + | </StructureSection> | |
| - | [[2zhs]], [[2zht]], [[2zhu]], [[2zhv]], [[1sgz]] - hBACE-1 catalytic domain – human<br /> | ||
| - | [[1w50]] - hBACE-1 catalytic domain (mutant)<br /> | ||
| - | [[3kyr]], [[3n4l]], [[3nsh]], [[3ixk]], [[3msj]], [[3msk]], [[3msl]], [[3msm]], [[3i25]], [[3k5d]], [[3k5f]], [[3k5g]], [[3l38]], [[3l3a]], [[3lhg]], [[3lnk]], [[3lpi]], [[3lpj]], [[3lpk]], [[2wjo]], [[3ind]], [[3ine]], [[3inf]], [[3inh]], [[3ixj]], [[3k5c]], [[3l58]], [[3l59]], [[3l5b]], [[3l5c]], [[3l5d]], [[3l5e]], [[3l5f]], [[3in3]], [[3in4]], [[3kmx]], [[3kmy]], [[3kn0]], [[3ivh]], [[3ivi]], [[3igb]], [[3h0b]], [[3duy]], [[3dv1]], [[3dv5]], [[2vkm]], [[3dm6]], [[2zdz]], [[2ze1]], [[2qu2]], [[2qu3]], [[3cib]], [[3cic]], [[3cid]], [[2qmg]], [[2qk5]], [[2qmd]], [[2qmf]], [[2qp8]], [[2p8h]], [[2q11]], [[2q15]], [[2p4j]], [[2p83]], [[2iqg]], [[2hiz]], [[2hm1]], [[3f3e]], [[3f3f]], [[2g94]], [[2fdp]], [[1ym2]], [[1ym4]], [[1xs7]], [[2f3e]], [[2f3f]], [[3ohf]], [[3ohh]], [[3pi5]], [[3qbh]], [[3s2o]], [[3ooz]], [[3r2f]], [[3s7l]], [[3s7m]], [[3skf]], [[3skg]], [[3u6a]] - hBACE-1 catalytic domain+inhibitor<br /> | ||
| - | [[2xfi]], [[2xfj]], [[2xfk]], [[3hvg]], [[3hw1]], [[2wez]], [[2wf0]], [[2wf1]], [[2wf2]], [[2wf3]], [[2wf4]], [[3exo]], [[2zjh]], [[2zji]], [[2zjk]], [[2zjl]], [[2zjm]], [[2zjn]], [[3fkt]], [[3ckp]], [[3ckr]], [[2vnm]], [[2vnn]], [[2qzk]], [[2ph8]], [[2qzl]], [[3bra]], [[3buf]], [[3bug]], [[3buh]], [[2vie]], [[2vij]], [[2viy]], [[2viz]], [[2vj6]], [[2vj7]], [[2vj9]], [[2ntr]], [[2va5]], [[2va6]], [[2va7]], [[2oah]], [[2ph6]], [[2oph]], [[2of0]], [[2ohk]], [[2ohl]], [[2ohm]], [[2ohn]], [[2ohq]], [[2ohr]], [[2ohs]], [[2oht]], [[2ohu]], [[2irz]], [[2is0]], [[2b8v]], [[2b8l]], [[1tqf]], [[1w51]], [[2ohp]], [[2zjj]], [[3rsv]], [[3rsx]], [[3rth]], [[3rtm]], [[3rtn]], [[3ru1]], [[3rvi]] - hBACE-1 catalytic domain (mutant)+inhibitor<br /> | ||
| - | [[2zhr]], [[1xn2]], [[1xn3]], [[1m4h]], [[1fkn]] - hBACE-1 catalytic domain + peptide inhibitor<br /> | ||
| - | [[3r1g]] - hBACE-1 catalytic domain + FAB<br /> | ||
| - | [[2ewy]] - hBACE-2 residues 16-398 + inhibitor | ||
==References== | ==References== | ||
| Line 77: | Line 71: | ||
*Wilquet V, Strooper BD (2004). "Amyloid-beta Precursor Protein Processing in Neurodegeneration". ''Current Opinion in Neurobiology'' '''14''':582-588. | *Wilquet V, Strooper BD (2004). "Amyloid-beta Precursor Protein Processing in Neurodegeneration". ''Current Opinion in Neurobiology'' '''14''':582-588. | ||
*Xu, Y., Li, M., Greenblatt, H.M., Chen, W., Paz, A., Dym, O., Peleg, Y., Chen, T., Shen, X., He, J., Jiang, H., Silman, I. & Sussman, J.L. (2012). “Flexibility of the flap in the active site of BACE1 as revealed by crystal structures and molecular dynamics simulations” Acta Crystallogr D Biol Crystallogr D68, 13-25. | *Xu, Y., Li, M., Greenblatt, H.M., Chen, W., Paz, A., Dym, O., Peleg, Y., Chen, T., Shen, X., He, J., Jiang, H., Silman, I. & Sussman, J.L. (2012). “Flexibility of the flap in the active site of BACE1 as revealed by crystal structures and molecular dynamics simulations” Acta Crystallogr D Biol Crystallogr D68, 13-25. | ||
| + | |||
| + | ==See also== | ||
| + | |||
| + | [[he:Beta Secretase (Hebrew)]] | ||
| + | [[ar:Beta Secretase (Arabic)]] | ||
[[Category:Topic Page]] | [[Category:Topic Page]] | ||
Current revision
| |||||||||||




References
- Alzheimer's Disease: Unraveling the Mystery. US Department of Health and Human Services, National Institute on Aging, NIH. 2008.
- CambridgeJournals. <http://journals.cambridge.org/fulltext_content/EPH/EPH87_04/S0958067001024046g003.htm>
- Patel S, Vuillard L, Cleasby A, Murray CW, Yon J (2004). "Apo and Inhibitor Complex Structures of BACE (β-secretase)". J.Mol.Biol. 343:407.
- Vassar R, Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA, Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, Luo Y, Fisher S, Fuller J, Edenson S, Lile J, Jarosinski MA, Biere AL, Curran E, Burgess T, Louis JC, Collins F, Treanor J, Rogers G, Citron M (1999). "β-Secretase Cleavage of Alzheimer's Amyloid Precursor Protein by the Transmembrane Aspartic Protease BACE". Science 286:735-741.
- Willem M, Dewachter I, Smyth N, Dooren TV, Borghgraef P, Haass C, Leuven FV (2004). "β-Site Amyloid Precursor Protein Cleaving Enzyme 1 Increases Amyloid Deposition in Brain Parenchyma but Reduces Cerebrovascular Amyloid Angiopathy in Aging BACE x APP[V717I] Double-Transgenic Mice". The American Journal of Pathology 165:1621-1631.
- Wilquet V, Strooper BD (2004). "Amyloid-beta Precursor Protein Processing in Neurodegeneration". Current Opinion in Neurobiology 14:582-588.
- Xu, Y., Li, M., Greenblatt, H.M., Chen, W., Paz, A., Dym, O., Peleg, Y., Chen, T., Shen, X., He, J., Jiang, H., Silman, I. & Sussman, J.L. (2012). “Flexibility of the flap in the active site of BACE1 as revealed by crystal structures and molecular dynamics simulations” Acta Crystallogr D Biol Crystallogr D68, 13-25.
See also
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, Alexander Berchansky, Daniel Santos, Jaime Prilusky, Joel L. Sussman, David Canner
