Tenebrio molitor Antifreeze Protein (TmAFP)
From Proteopedia
(Difference between revisions)
| (95 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
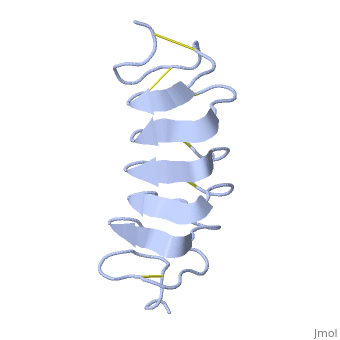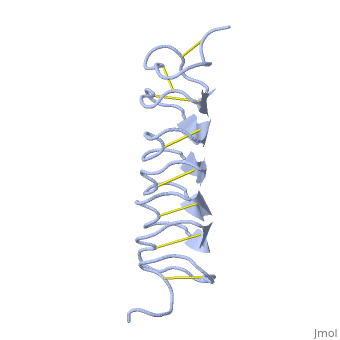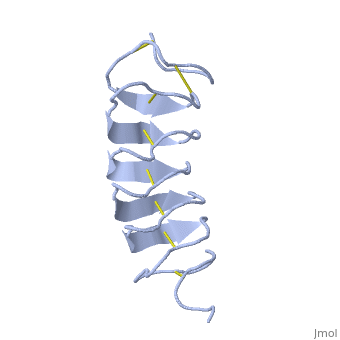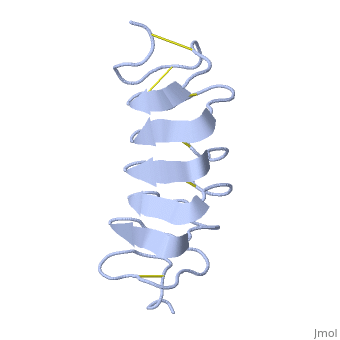
| - | <StructureSection load='1l1i' size=' | + | <StructureSection load='1l1i' size='400' side='right' caption='''Tenebrio Molitor'' antifreeze protein NMR structure (PDB code [[1l1i]]).' scene=''> |
__TOC__ | __TOC__ | ||
== Introduction == | == Introduction == | ||
| + | Antifreeze proteins (AFPs) are produced by certain vertebrates, plants, fungi and bacteria that permit their survival in subzero environments. AFPs bind to ice crystals to inhibit growth of ice that would otherwise be fatal to those organisms. | ||
| + | AFPs are classified into two groups on the basis of their activity (Thermal Hysteresis or TH, see below in Function), moderate and hyperactive AFPs. | ||
| + | Moderate AFP's usually originate from fish that live in the polar area in which the water temperature can be low as -2 degrees Celsius . | ||
| + | Hyperactive AFP usually originate from insects (but recently have been discovered in other organisms including bacteria) that live in areas where the temperature is much lower and can be as low as -40 degrees Celsius. | ||
| + | <ref>DOI doi:10.1016/j.cryobiol.2006.06.006</ref> | ||
| - | ''Tm''AFP is | + | ''Tm''AFP is a hyperactive antifreeze protein and its origin is the Tenebrio Molitor beetle (Mealworm). These beetles are found in dark and very cold areas such as the polar region. Due to ''Tm''AFP, the Tenebrio Molitor beetle is freezing tolerant which provides protection against physical and osmotic stresses. ''Tm''AFP is shaped like a slightly flattened cylinder 32Å in length, and 6.5X 14Å in the pseudo,rectangular cross section and is one of the smallest Beta Hellix AFP known to date. |
| - | + | ||
| + | This is a <scene name='61/612804/Rainbow/1'>Rainbow</scene> view of the protein presenting the N terminal in blue and the C terminal in red. | ||
== Structure == | == Structure == | ||
The protein consists of 84 amino acids and the molecular weight is 8.4 kDA. | The protein consists of 84 amino acids and the molecular weight is 8.4 kDA. | ||
| - | ''Tm''AFP is protein rich of | + | ''Tm''AFP is protein rich of Threonine and Cystein and forms a regular parallel beta-helix. It is composed of 7 tandem repeats which consist of 12 amino acids (TCTxSxxCxxAx). <scene name='61/612804/Tct/1'>TCT </scene> or ACT (<span style="color:purple;background-color:white;font-weight:bold;">Thr</span>, <span style="color:aqua;background-color:white;font-weight:bold;">Cys</span>, <span style="color:purple;background-color:white;font-weight:bold;">Thr</span>) motifs are aligned to form a <scene name='61/612804/Beta_sheet/2'>flat Beta sheet</scene> along one side of the molecule the Beta sheets right handed which are the binding site of the protein.<ref name="one">DOI 10.1529/biophysj.104.051169</ref> The rest of the tandem repeat forms the loop which enables a very organized structure of the protein. Cys lockets systematically along the tandem repeats, are paired to provide the <scene name='61/612804/Cys/1'>disulphide bonds</scene> which contribute significantly to the stability of the protein. <scene name='61/612804/Cys/2'>Six</scene> of the eight disulphide bounds construct a near perfect alignment yielding a structure that allows binding to the ice lattice. The other <scene name='61/612804/2disulphide/1'>two disulphide bonds</scene> in the N-terminal region do not fit this pattern and are involved in a capping structure. The extraordinary tightness of the 12 amino-acids turn is also facilitated by intra-loop <scene name='61/612804/Hbond/2'>hydrogen bond</scene> connections between backbone CO and NH groups. |
| - | TCT ( | + | [[Image:406322aa.2.jpg|230px|right]] |
| - | The rest of the tandem repeat forms the loop which enables very organized structure of the protein. | + | ''Tm''AFP is the one of smallest Beta-Helix with only 12 amino acids per turn. Therefore, it has a very narrow bore, which is constricted and further bisected by disulphide bonds to form two channels, leaving no room for hydrophobic core. The few Hydrophobic residues <scene name='61/612804/Hydrophobic/2'>Val25, Val34, Phe58 and Tyr70</scene> have their side chains projecting outwards to the solvent. In the core there is room only for the relatively small side chains of the conserves Ser and Ala which project into the core, on either side of the bisecting disulphide bridges.<ref name="two">DOI 10.1038/35018604</ref> |
| - | + | ||
| - | + | ||
| - | + | == Ice binding site == | |
| - | ''Tm''AFP is the smallest Beta-Helix with only 12 amino acids per turn. Therefore, it has a very narrow bore, which is constricted and further bisected by disulphide bonds to form two channels, leaving no room for hydrophobic core. | ||
| - | The few Hydrophobic residues <scene name='61/612804/Hydrophobic/2'>val25, val34, phe58, tyr70</scene> have their side chains projecting outwards to the solvent. In the core there is room only for the relatively small side chains of the conserves Serine and Alanine to project into the core, on either side of the bisecting disulphide bridge.<ref>DOI 10.1038/35018604</ref> | ||
| - | + | [[Image:406322ad.2.jpg|230px|right]] | |
| + | The two dimensional arrays of <scene name='61/612804/Thr/3'>Thr</scene> side chain makes a remarkably good match to the repeated spacing between oxygen atoms in the ice lattice on the prism plane, and a reasonable match to the basal plane, This is reason why the activity of ''Tm''AFP ( Thermal hysteresis) is much higher than the activity of AFP from Fish (6 degrees Celcius and 1 degree celcius respectively)<ref>doi:10.1016/S0968-0004(01)02028-X</ref> | ||
| - | == Ice binding site == | ||
| + | '''Lattice matching model for TmAFP binding to ice:''' | ||
| + | a- Prism plane. | ||
| + | |||
| + | b- Basal plane. | ||
| + | |||
| + | |||
| + | |||
| + | |||
| + | |||
| + | In solution the protein is monomeric and it can bind to ice just in this formation. The protein crystallized as a <scene name='61/612804/Dimer/1'>dimer</scene>, the dimerization occurs along the surface of the beta sheets. The 2 units of the dimer do not directly interact with each other, the contact between them is mediated by highly ordered ranks of water which form hydrogen bonding with Threonine residue. Each water molecule forms two hydrogen bonds to the closer monomer and one to the distant monomer. The distance between two adjacent waters is 4.64±0.20Å the same distance as between Threonine residues 4.64±0.23Å. A good two dimensional match to the ice lattice including all 3 ranks of oxygen atoms (2 from Thr and 1 from water), implying that the ordered water molecules cold act as part of an ice surface and directly participate in the AFP-ice interaction. The regular array formed by this 3 ranks of oxygen atoms can be seen as a small piece of one layer thick ice to be incorporated into a large ice lattice. There is no need to readjust Threonine side chains, because they don’t present steric interference <ref name="two">DOI 10.1038/35018604</ref>. The relevance of the dimer is to show the similarity of the binding of monomer to ice and the binding of the dimer. Dimer binds through Water molecules mimic the same way as the monomer binds to ice. [[Image:406322ac.2.jpg|300px|right]] | ||
| + | |||
| + | |||
| + | '''Dimer of TmAFP and organization of external water:''' | ||
| - | + | a- A TmAFP pair dimerized through hydrogen bonding to two ranks of ordered waters. | |
| - | + | b- Surface presentation of TmAFP with the closest rank of water molecules. | |
| - | + | ||
| - | The 2 units of the dimer do not directly interact with each other, the contact between them is mediated by highly ordered ranks of water which form hydrogen bonding with Threonine residue. | ||
| - | Each water molecule forms two hydrogen bonds to the closer monomer and one to the distant monomer. | ||
| - | The distance between two adjacent waters is 4.64±0.20Å the same distance as between Threonine residues 4.64±0.23Å. | ||
| - | A good two dimensional match to the ice lattice including all 3 ranks of oxygen atoms (2 from Thr and 1 from water), implying that the ordered water molecules cold act as part of an ice surface and directly participate in the AFP-ice interaction. | ||
| - | The regular array formed by this 3 ranks of oxygen atoms can be seen as a small piece of one layer thick ice to be incorporated into a large ice lattice. | ||
| - | + | == Function == | |
| + | The function of the ''Tm''AFP (and other Antifreeze protein) is TH. TH activity is due to an adsorption inhibition mechanism that’s states that AFPs binds to ice surface and allow ice crystal growths only in surface regions between the bound AFP.[[Image:Unnamed.jpg|250px|right]] Growing curvature causes an increase in surface energy, making the transformation of water into ice lass energetically favorable. Because of that the AFPs lower freezing temperature below the melting point. The difference between the melting and freezing point of the ice called thermal hysteresis activity. | ||
| - | == Relevance == | ||
| - | == | + | == The difference between TmAFP (hyperactive AFP) and Type I AFP (moderate AFP) == |
| + | TmAFP has a special structure of a short very regular beta helix, which yields a binding site consisting of a two dimensional surface: <scene name='61/612804/Thr/3'>two arrays of Thr residues</scene> that can bind to the two planes of ice i.e. prism plane and basal plane. This is why the TH activity can reach 6 degrees Celsius in Hyperactive protein. | ||
| + | In contrast <scene name='61/612804/Afp1/1'>Type I AFP</scene>, from the fish winter flounder (moderate AFP are usually fish origin), structure is alpha helix allows just one dimensional surface of Thr residue and thus can bind only to one plane of ice (the prism plan). Because of that The TH activity is much lower, maximum TH activity of moderate AFP is 1 degree Celsius. | ||
| - | ==3D structures of antifreeze protein== | ||
[[Antifreeze protein]] | [[Antifreeze protein]] | ||
Current revision
| |||||||||||




References
- ↑ Scotter AJ, Marshall CB, Graham LA, Gilbert JA, Garnham CP, Davies PL. The basis for hyperactivity of antifreeze proteins. Cryobiology. 2006 Oct;53(2):229-39. Epub 2006 Aug 2. PMID:16887111 doi:http://dx.doi.org/10.1016/j.cryobiol.2006.06.006
- ↑ Liu K, Jia Z, Chen G, Tung C, Liu R. Systematic size study of an insect antifreeze protein and its interaction with ice. Biophys J. 2005 Feb;88(2):953-8. PMID:15713600 doi:http://dx.doi.org/10.1529/biophysj.104.051169
- ↑ 3.0 3.1 Liou YC, Tocilj A, Davies PL, Jia Z. Mimicry of ice structure by surface hydroxyls and water of a beta-helix antifreeze protein. Nature. 2000 Jul 20;406(6793):322-4. PMID:10917536 doi:10.1038/35018604
- ↑ doi: https://dx.doi.org/10.1016/S0968-0004(01)02028-X
