|
|
| (38 intermediate revisions not shown.) |
| Line 1: |
Line 1: |
| | + | <StructureSection load='1iwg' size='450' side='right' scene='Insert optional scene name here' caption='AcrB transporter ([[1iwg]])' >__NOTOC__ |
| | {{UniHelsinki_ProteinCourse_2015}} | | {{UniHelsinki_ProteinCourse_2015}} |
| - | <Structure load='1IWG' size='350' frame='true' align='right' caption='AcrB' scene='Insert optional scene name here' /><!-- PLEASE ADD YOUR CONTENT BELOW HERE --> | |
| | | | |
| - | == Introduction == | |
| | | | |
| - | [[Image:efflux pump.jpg|thumb|left|200px|Hypothetical structure of tripartite efflux pumps in Gram-negative bacteria, e.g. the AcrA/AcrB/TolC efflux pump from ''E. coli'']]
| + | == '''AcrB Transporter''' == |
| | | | |
| - | In Gram-negative bacteria, such as ''Escherichia coli'', one of the main reasons behind bacterial multidrug resistance is resistance nodulation cell division (RND) transporters which pumps a wide range of antibiotics, dyes, bile salts and detergents out of the cell by proton motive force. Naturally, AcrB is speculated to function by pumping out of bile salts and their derivatives from the natural habitat of ''E. coli''. for their survival with the presence of the high concentrations of these detergents. AcrB is the major RND transporter in ''E. coli''. Three proteins form the tripartite multidrug efflux system that pumps the drugs out from the cell. The AcrB transporter, the inner membrane component of the system, cooperates with two other proteins: membrane fusion protein AcrA and an outer membrane channel TolC. The structure of the complex indicates that the drugs are transported out of the cell in a three-step procedure. [1, 2, 3]
| + | [[Image:efflux pump.jpg|thumb|left|200px|Figure 1. Hypothetical structure of tripartite efflux pumps in Gram-negative bacteria, e.g. the AcrA/AcrB/TolC efflux pump from ''E. coli''<ref name= "Nakashima"/>]] |
| | | | |
| - | <scene name='70/700000/Acrb_transporter/1'>TextToBeDisplayed</scene> | + | In Gram-negative bacteria, such as Escherichia coli, one of the main reasons behind bacterial multidrug resistance is resistance nodulation cell division (RND) transporters which pump a wide range of dyes, bile salts, detergents and structurally unrelated antibiotics out of the cell by proton motive force driven efflux. The natural function of the efflux system is currently under debate. AcrB is the major RND transporter in E. coli. Three proteins form the tripartite multidrug efflux system that pumps the drugs out from the cell. The AcrB transporter, the inner membrane component of the system, cooperates with two other proteins: membrane fusion protein AcrA and an outer membrane channel TolC. The TolC component is connected to outer membrane and AcrB is connected to the inner membrane. The AcrA connects TolC and AcrB together (Fig 1). The structure of the complex indicates that the drugs are transported out of the cell in a three-step procedure.<ref name= "Nakashima">PMID: 12607261</ref><ref name= "Klaas">PMID: 19166984</ref><ref name= "Murasaki">PMID: 18644451</ref> |
| | | | |
| | | | |
| - | This is a default text for your page ''''''. Click above on '''edit this page''' to modify. Be careful with the < and > signs. | |
| - | You may include any references to papers as in: the use of JSmol in Proteopedia <ref>DOI 10.1002/ijch.201300024</ref> or to the article describing Jmol <ref>PMID:21638687</ref> to the rescue. | |
| | | | |
| | | | |
| - | == Structure == | |
| | | | |
| - | The tripartite multidrug efflux system contains three components: AcrB, the inner membrane component, AcrA, the membrane fusion protein and TolC, the outer membrane channel. The TolC component is connected to outer membrane and AcrB is connected to the inner membrane. The AcrA connects TolC and AcrB together (Fig 1.). In the AcrB component there is three domains: TolC docking domain, pore domain and the transmembrane domain. The AcrB consists of 1 049 amino acids and the three protomers of AcrB are organized as a homo<scene name='70/700000/Acrb_trimer/1'>trimer</scene> (Fig. 2). The appearance of the trimer is of a jellyfish. The N- and C-terminal half’s show similar structural architecture and indicates at early gene duplication events. The trimer, formed by AcrB monomers, appears to be stabilized by the intermonomer connecting loops. [1, 2] | |
| | | | |
| - | [[Image:Nature01050-f2.2.jpg|thumb|right|350px|text]]
| + | == Structure == |
| - | '''Transmembrane domain structure'''
| + | |
| | | | |
| - | Twelve α-helices of each promoter forms the transmembrane domain. Six α-helices in the N-terminal and the six in C-terminal are arranged symmetrically. These helices are long and they reach outside the cytoplasmic surface of the membrane. The membrane domain contains an additional extra-membrane a-helix (Ia) located between <scene name='70/700000/Acrb_tm_6/2'>TM6</scene> and TM7 attached to the cytoplasmic membrane surface (Fig. 3a and b). Amino acid residues 860-868 (TM8) are in a disordered state. Between TM8 and TM7 locates a groove within the transmembrane domain of each promoter. Through the disordered region of the top of TM8, the groove is connected with the cavity. This domain contains three functionally critical residues: Asp407, Asp480 and Lys940. When these are mutated the whole complex loses its drug resistance.
| + | [[Image:Nature01050-f2.2.jpg|thumb|left|200px|Figure 2. Structure of AcrB. a) Side view of the ribbon representation. The three monomers are |
| | + | individually coloured (blue, green and red), b) Top view of a ribbon representation, c) Structure within a slab of the transmembrane domain parallel to the membrane plane near the periplasmic surface.<ref name= "Nakashima"/>]] |
| | + | [[Image:Figure5.png|thumb|right|600px|Figure 3. The structure of a single monomer. A) Topology diagram of the protomer, B) A stereo view of ribbon representation of a protomer viewed from outside the pore domain<ref name= "Nakashima"/>]] |
| | + | [[Image:Fig groove.png|thumb|left|300px|Figure 4. The groove in the transmembrane domain highlighted in green<ref name= "Nakashima"/>]] |
| | | | |
| | + | In the AcrB, there is three domains: TolC docking domain, pore domain and the transmembrane domain. The AcrB consists of 1 049 amino acids and the three monomers of AcrB are organized as a trimer (Fig. 2). The first structure of the trimer was derived in symmetric manner <scene name='70/700000/Acrb_trimer/1'>trimer</scene> (Fig. 2) at 3.5 Å resolution. Later, a new 2.9 Å asymmetric conformation of trimeric AcrB was obtained and three structurally different monomers were spotted<ref>PMID: 16946072</ref>. The appearance of the trimer is of a jellyfish. The N- and C-terminal half’s show similar structural architecture and indicates at early gene duplication events. The trimer, formed by AcrB monomers, appears to be stabilized by the intermonomer connecting loops.<ref name= "Nakashima"/> |
| | | | |
| - | '''Pore domain structure''' | |
| | | | |
| - | The pore domain consists of subdomains <scene name='70/700000/Acrb_pn1/1'>PN1</scene>, <scene name='70/700000/Acrb_pn2/1'>PN2</scene>, <scene name='70/700000/Acrb_pc1/1'>PC1</scene> and <scene name='70/700000/Acrb_pc2/1'>PC2</scene>. These subdomains have a characteristic structural motif: two b-strand–a-helix–b-strand motifs are directly repeated and sandwiched with each other. This motif forms a structure in which two a-helices are located on a four-stranded antiparallel b-sheet. Three α-helices forms a pore in the middle of the structure. The headpiece of the structure (the TolC docking domain) forms a funnel like structure in the middle of the domain. The pore connects with the bottom of the funnel (Fig 2B.). The α-helices that forms the pore are from the PN1 subunit (Fig. 2C). Between PN2 and PC2, there are vestibules open at the side of the headpiece into the periplasm. They have access to a cavity at the bottom of the pore. Analysis with the AcrA has suggested that PC1 and PC2 subdomains has a role in attaching the AcrA to the complex. Analyses has suggested that C-terminal domain residues 290-357 has a main role in interacting with AcrA. [1, 2]
| + | '''<scene name='70/700000/Tolc_docking/1'>TolC docking domain</scene>''' |
| | | | |
| | + | The TolC docking domain, located in the periplasm, has two subdomains: <scene name='70/700000/Acrb_dn/1'>DN</scene> and <scene name='70/700000/Acrb_dc/1'>DC</scene>. The subdomains contain a four-stranded mixed β-sheet. The TolC domain forms a funnel-like structure that has a similar diameter as the bottom of TolC. The TolC docking domain of AcrB and the bottom of TolC fits well with each other for connecting.<ref name= "Nakashima"/> |
| | | | |
| - | '''TolC docking domain structure''' | |
| | | | |
| - | The TolC docking domain has two subdomains: <scene name='70/700000/Acrb_dn/1'>DN</scene> and <scene name='70/700000/Acrb_dc/1'>DC</scene>. The subdomains contains a four-stranded mixed b-sheet. The domain forms a funnel-like structure that has a similar diameter as the bottom of TolC. The TolC docking domain of AcrB and the bottom of TolC fits well with each other for connecting. [1]
| + | '''<scene name='70/700000/Pore_domain/1'>Pore domain </scene>''' |
| | | | |
| | + | The pore domain consists of subdomains <scene name='70/700000/Acrb_pn1/1'>PN1</scene>, <scene name='70/700000/Acrb_pn2/1'>PN2</scene>, <scene name='70/700000/Acrb_pc1/1'>PC1</scene> and <scene name='70/700000/Acrb_pc2/1'>PC2</scene> (Fig. 3A). These subdomains have a characteristic structural motif: two β-strand–α-helix–β-strand motifs are directly repeated and sandwiched with each other. This motif forms a structure in which two α-helices are located on a four-stranded antiparallel β-sheet. Three α-helices from each PN1 subdomains form a pore in the middle of the structure (Fig. 3B). The pore connects with the bottom of the funnel-like structure of the TolC docking domain. The extramembrane part of the central membrane hole, namely, the central cavity is present at the proximal end of the pore (Fig. 2c). Between PN2 and PC2, there are vestibules open at the side of the pore domain into the periplasm (Fig. 3B). They have access to the central cavity. Analysis with the AcrA has suggested that PC1 and PC2 subdomains are of great importance in attaching the AcrA to the complex. Studies have suggested that C-terminal domain residues <scene name='70/700000/Acrb_290-357/1'>290-357</scene> play a major role in interacting with AcrA.<ref name= "Nakashima"/><ref name= "Klaas"/> |
| | | | |
| - | == Function == | |
| | | | |
| - | [[Image:Nature01050-f5.2.jpg|thumb|left|350px|text]] | |
| | | | |
| - | Possible mechanism of transport function has been postulated. ArcB can cooperate with TolC in the TolC docking domain forming a direct pathway from the cytoplasm to the extracellular milieu. The end of the headpiece is the narrow funnel connected with the pore, whose end is connected with the extramembrane part of the central membrane hole, namely, the cavity. There might be two pathways for the substrate translocation to the cavity. One is the groove located between <scene name='70/700000/Acrb_tm8/3'>TM8</scene> and <scene name='70/700000/Acrb_tm7/2'>TM7</scene> in the transmembrane domain of each promoter (Fig 1C). The other is the vestibules which are open at the side of the headpiece between PN2 and PC2 of the pore domains. The substrates located in the inner leaflet of the membrane and cytoplasm would get access to the cavity through the transmembrane groove, while the substrates which are on the outer space or in the outer leaflet of the membrane are more likely to be transported to the cavity through the vestibules.
| + | '''<scene name='70/700000/Transmembrane_domain/2'>Transmembrane domain</scene>''' |
| - | Studies have been done on the substrate binding and specificity and the periplasmic part of the tripartite efflux system is found important to the substrate specificity. In the study on antibiotics to AcrB, Phe 386 (TM3) was reported as one the main hydrophobic contacts. However, currently, the theoretical explanation of the wide variety of substrates is still lacking.
| + | |
| - | Proton-motive force has been proposed to be coupled with the tripartite efflux system. It is the transmembrane domain where binding and release of protons happen. Certain key residues had been identified as crucial to the protons translocation. They are the residues Lys940 (<scene name='70/700000/Acrb_tm10/2'>TM10</scene>) and Asp407 and 408 (TM4) harbored in <scene name='70/700000/Acrb_tm_4/2'>TM4</scene> and TM10 in each protomer. When the ion pairs between them are disrupted due to the transient protonation of residues mentioned above, there may be conformational change of TM4 and TM10. Through possible remote conformational coupling, the conformational change of TM4 and TM10 may induce the opening of the pore.
| + | |
| | | | |
| | + | Twelve α-helices of each monomer forms the transmembrane domain (TM1-12). Six α-helices in the N-terminal and the six in C-terminal are arranged symmetrically. These helices are long and they reach outside the cytoplasmic surface of the membrane. There is an α-helix (Iα) located between TM6 and TM7 in the transmembrane domain. This Iα attaches to the cytoplasmic membrane surface. Between <scene name='70/700000/Acrb_tm8/3'>TM8</scene> and <scene name='70/700000/Acrb_tm7/2'>TM7</scene> locates a groove within the transmembrane domain of each monomer. Amino acid residues 860-868 in TM8 are in a disordered state. Through the disordered region of the top of TM8, the groove is connected with the cavity. This domain contains three functionally critical residues: Asp407, Asp408 and Lys940. The residues are shown in this <scene name='70/700000/Acrb_residues/2'>link</scene>, Asp407 as pink, Asp408 as green and Lys940 as blue. When these are mutated the whole complex loses its drug resistance.<ref name= "Nakashima"/> |
| | | | |
| | | | |
| | + | == Function == |
| | + | |
| | + | [[Image:Nature01050-f5.2.jpg|thumb|right|350px|Figure 4. Proposed model of the AcrB/AcrA/TolC complex and the schematic mechanism of multidrug export mediated by AcrAB/TolC system<ref name= "Nakashima"/>]] |
| | | | |
| | + | Possible mechanism of transport function has been postulated. ArcB can cooperate with TolC in the TolC docking domain forming a direct pathway from the cytoplasm to the extracellular milieu. Based on the information of structure, there might be two pathways for the substrate translocation to the central cavity. One is the groove located between TM8 and TM7 in the transmembrane domain of each monoter (Fig 1C). The other is the vestibules which are between PN2 and PC2 of the pore domains. The substrates located in the inner leaflet of the membrane and cytoplasm would get access to the central cavity through the transmembrane groove, while the substrates that are on the outer space or in the outer leaflet of the membrane are more likely to be transported to the cavity through the vestibules.<ref name= "Nakashima"/> |
| | + | Studies have been done on the substrate specificity and the periplasmic part of the tripartite efflux system is found important to the substrate specificity. In the study on antibiotics to AcrB, Phe<scene name='70/700000/Acrb_386/1'>386</scene> (TM3) was reported as one the main hydrophobic contacts. However, currently, the theoretical explanation of the wide variety of substrates is still lacking.<ref name= "Nakashima"/> |
| | + | The tripartite efflux system coupled with proton-motive force across the cytoplasmic membrane. It is the binding and release of protons in the transmembrane domain that is crucial to the energy transduction.<ref name= "Klaas"/> Certain key residues had been identified as crucial to the protons translocation. They are the residues Lys940 (TM10) and Asp407 and 408 (TM4) harbored in <scene name='70/700000/Acrb_tm_4/2'>TM4</scene> and <scene name='70/700000/Acrb_tm10/2'>TM10</scene> in each monomer. When the ion pairs between them are disrupted due to the transient protonation of residues mentioned above, there may be conformational change of TM 4 and TM10. Through possible remote conformational coupling, the conformational change of TM 4 and TM10 may induce the opening of the pore.<ref name= "Nakashima"/> |
| | | | |
| | | | |
| Line 55: |
Line 56: |
| | | | |
| | | | |
| - | == Relevance == | |
| | | | |
| - | [[Image:a5fbdffa56.jpg]] | |
| | | | |
| - | == Structural highlights == | |
| | | | |
| - | This is a sample scene created with SAT to <scene name="/12/3456/Sample/1">color</scene> by Group, and another to make <scene name="/12/3456/Sample/2">a transparent representation</scene> of the protein. You can make your own scenes on SAT starting from scratch or loading and editing one of these sample scenes. | |
| | | | |
| - | </StructureSection> | |
| | == References == | | == References == |
| | <references/> | | <references/> |
|
| This Sandbox is Reserved from 15/04/2015, through 15/06/2015 for use in the course "Protein structure, function and folding" taught by Taru Meri at the University of Helsinki. This reservation includes Sandbox Reserved 1081 through Sandbox Reserved 1090.
|
To get started:
- Click the edit this page tab at the top. Save the page after each step, then edit it again.
- Click the 3D button (when editing, above the wikitext box) to insert Jmol.
- show the Scene authoring tools, create a molecular scene, and save it. Copy the green link into the page.
- Add a description of your scene. Use the buttons above the wikitext box for bold, italics, links, headlines, etc.
More help: Help:Editing
|
AcrB Transporter
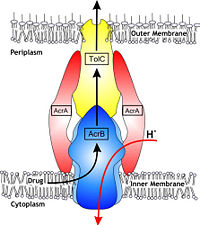 Figure 1. Hypothetical structure of tripartite efflux pumps in Gram-negative bacteria, e.g. the AcrA/AcrB/TolC efflux pump from E. coli[1]In Gram-negative bacteria, such as Escherichia coli, one of the main reasons behind bacterial multidrug resistance is resistance nodulation cell division (RND) transporters which pump a wide range of dyes, bile salts, detergents and structurally unrelated antibiotics out of the cell by proton motive force driven efflux. The natural function of the efflux system is currently under debate. AcrB is the major RND transporter in E. coli. Three proteins form the tripartite multidrug efflux system that pumps the drugs out from the cell. The AcrB transporter, the inner membrane component of the system, cooperates with two other proteins: membrane fusion protein AcrA and an outer membrane channel TolC. The TolC component is connected to outer membrane and AcrB is connected to the inner membrane. The AcrA connects TolC and AcrB together (Fig 1). The structure of the complex indicates that the drugs are transported out of the cell in a three-step procedure.[1][2][3]
Structure
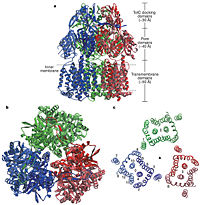 Figure 2. Structure of AcrB. a) Side view of the ribbon representation. The three monomers are individually coloured (blue, green and red), b) Top view of a ribbon representation, c) Structure within a slab of the transmembrane domain parallel to the membrane plane near the periplasmic surface. [1]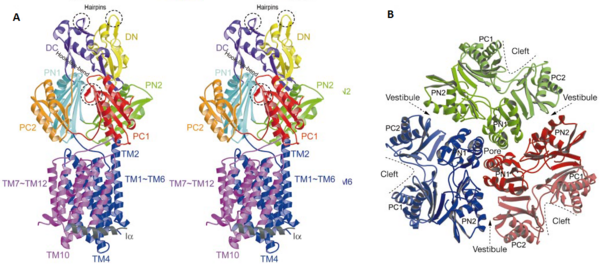 Figure 3. The structure of a single monomer. A) Topology diagram of the protomer, B) A stereo view of ribbon representation of a protomer viewed from outside the pore domain [1]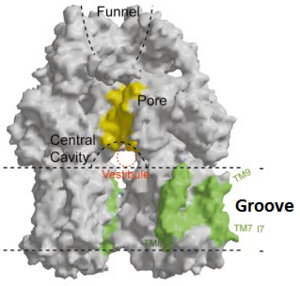 Figure 4. The groove in the transmembrane domain highlighted in green [1]In the AcrB, there is three domains: TolC docking domain, pore domain and the transmembrane domain. The AcrB consists of 1 049 amino acids and the three monomers of AcrB are organized as a trimer (Fig. 2). The first structure of the trimer was derived in symmetric manner (Fig. 2) at 3.5 Å resolution. Later, a new 2.9 Å asymmetric conformation of trimeric AcrB was obtained and three structurally different monomers were spotted[4]. The appearance of the trimer is of a jellyfish. The N- and C-terminal half’s show similar structural architecture and indicates at early gene duplication events. The trimer, formed by AcrB monomers, appears to be stabilized by the intermonomer connecting loops.[1]
The TolC docking domain, located in the periplasm, has two subdomains: and . The subdomains contain a four-stranded mixed β-sheet. The TolC domain forms a funnel-like structure that has a similar diameter as the bottom of TolC. The TolC docking domain of AcrB and the bottom of TolC fits well with each other for connecting.[1]
The pore domain consists of subdomains , , and (Fig. 3A). These subdomains have a characteristic structural motif: two β-strand–α-helix–β-strand motifs are directly repeated and sandwiched with each other. This motif forms a structure in which two α-helices are located on a four-stranded antiparallel β-sheet. Three α-helices from each PN1 subdomains form a pore in the middle of the structure (Fig. 3B). The pore connects with the bottom of the funnel-like structure of the TolC docking domain. The extramembrane part of the central membrane hole, namely, the central cavity is present at the proximal end of the pore (Fig. 2c). Between PN2 and PC2, there are vestibules open at the side of the pore domain into the periplasm (Fig. 3B). They have access to the central cavity. Analysis with the AcrA has suggested that PC1 and PC2 subdomains are of great importance in attaching the AcrA to the complex. Studies have suggested that C-terminal domain residues play a major role in interacting with AcrA.[1][2]
Twelve α-helices of each monomer forms the transmembrane domain (TM1-12). Six α-helices in the N-terminal and the six in C-terminal are arranged symmetrically. These helices are long and they reach outside the cytoplasmic surface of the membrane. There is an α-helix (Iα) located between TM6 and TM7 in the transmembrane domain. This Iα attaches to the cytoplasmic membrane surface. Between and locates a groove within the transmembrane domain of each monomer. Amino acid residues 860-868 in TM8 are in a disordered state. Through the disordered region of the top of TM8, the groove is connected with the cavity. This domain contains three functionally critical residues: Asp407, Asp408 and Lys940. The residues are shown in this , Asp407 as pink, Asp408 as green and Lys940 as blue. When these are mutated the whole complex loses its drug resistance.[1]
Function
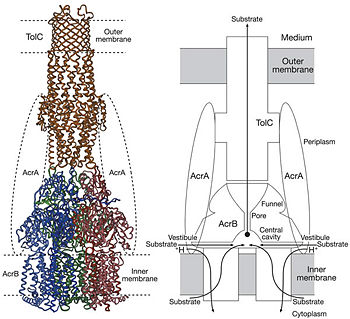 Figure 4. Proposed model of the AcrB/AcrA/TolC complex and the schematic mechanism of multidrug export mediated by AcrAB/TolC system [1]Possible mechanism of transport function has been postulated. ArcB can cooperate with TolC in the TolC docking domain forming a direct pathway from the cytoplasm to the extracellular milieu. Based on the information of structure, there might be two pathways for the substrate translocation to the central cavity. One is the groove located between TM8 and TM7 in the transmembrane domain of each monoter (Fig 1C). The other is the vestibules which are between PN2 and PC2 of the pore domains. The substrates located in the inner leaflet of the membrane and cytoplasm would get access to the central cavity through the transmembrane groove, while the substrates that are on the outer space or in the outer leaflet of the membrane are more likely to be transported to the cavity through the vestibules.[1]
Studies have been done on the substrate specificity and the periplasmic part of the tripartite efflux system is found important to the substrate specificity. In the study on antibiotics to AcrB, Phe (TM3) was reported as one the main hydrophobic contacts. However, currently, the theoretical explanation of the wide variety of substrates is still lacking.[1]
The tripartite efflux system coupled with proton-motive force across the cytoplasmic membrane. It is the binding and release of protons in the transmembrane domain that is crucial to the energy transduction.[2] Certain key residues had been identified as crucial to the protons translocation. They are the residues Lys940 (TM10) and Asp407 and 408 (TM4) harbored in and in each monomer. When the ion pairs between them are disrupted due to the transient protonation of residues mentioned above, there may be conformational change of TM 4 and TM10. Through possible remote conformational coupling, the conformational change of TM 4 and TM10 may induce the opening of the pore.[1]
References
- ↑ 1.00 1.01 1.02 1.03 1.04 1.05 1.06 1.07 1.08 1.09 1.10 1.11 1.12 Murakami S, Yamaguchi A. [Crystal structure of bacterial multidrug efflux transporter AcrB]. Tanpakushitsu Kakusan Koso. 2003 Jan;48(1):26-32. PMID:12607261
- ↑ 2.0 2.1 2.2 Pos KM. Drug transport mechanism of the AcrB efflux pump. Biochim Biophys Acta. 2009 May;1794(5):782-93. doi: 10.1016/j.bbapap.2008.12.015., Epub 2009 Jan 3. PMID:19166984 doi:http://dx.doi.org/10.1016/j.bbapap.2008.12.015
- ↑ Murakami S. Multidrug efflux transporter, AcrB--the pumping mechanism. Curr Opin Struct Biol. 2008 Aug;18(4):459-65. doi: 10.1016/j.sbi.2008.06.007., Epub 2008 Aug 9. PMID:18644451 doi:http://dx.doi.org/10.1016/j.sbi.2008.06.007
- ↑ Seeger MA, Schiefner A, Eicher T, Verrey F, Diederichs K, Pos KM. Structural asymmetry of AcrB trimer suggests a peristaltic pump mechanism. Science. 2006 Sep 1;313(5791):1295-8. PMID:16946072 doi:313/5791/1295
|





