|
|
| (119 intermediate revisions not shown.) |
| Line 1: |
Line 1: |
| | == '''Caspase-6 in ''Homo sapiens''' == | | == '''Caspase-6 in ''Homo sapiens''' == |
| | | | |
| - | <StructureSection load='4FXO' size='340' side='right' caption='Caspase-6' scene=''> | + | <StructureSection load='4IYR' size='340' side=http://proteopedia.org/wiki/index.php?title=User:Luke_Edward_Severinac/Sandbox_1&action=edit'right' caption='Caspase-6' scene=''> |
| - | Found at high concentrations in the brain and bordering tissues, Caspase-6 has been implicated in several neurological diseases including Alzheimer's and dementia[http://www.alz.org/]<ref name="ActiveRegofCasp6andNDdisease">PMID: 25340928 </ref>. It's primarily involved in apoptosis through a largely ambiguous mechanism. It is classified as an [https://en.wikipedia.org/wiki/Endopeptidase]endopeptidase as it cleaves an internal peptide bond of its substrate. It has relatively low specificity in the binding site which allows for a variety of substrates, including other caspase enzymes and neuronal proteins to bind<ref name="ZincMediatedCasp6">PMID: 22891250 </ref>. Furthermore, it is a part of the cysteine-aspartate family[https://en.wikipedia.org/wiki/Caspase], which have these critical amino acid residues in the active site of the enzyme. Caspase-6 has both an inactive zinc-bound conformation and an active ligand-bound conformation, which are largely regulated by variations in zinc concentration<ref name="ZincMediatedCasp6">PMID: 22891250 </ref>.
| + | Caspase-6 is an [https://en.wikipedia.org/wiki/Endopeptidase endopeptidase] involved in apoptosis. In terms of its catalytic function, it is a part of the [https://en.wikipedia.org/wiki/Caspase cysteine-aspartate family]. Before Caspase-6 becomes functional, the enzyme exists as a <scene name='75/752344/Caspase-6_zymogen/1'>procaspase</scene>, also known as a [https://en.wikipedia.org/wiki/Zymogen zymogen]. This zymogen exists as a <scene name='75/752344/Caspase-6_zymogen/1'>homodimer</scene>, whose <scene name='75/752344/Caspase-6_zymogen_realller/1'>monomeric units</scene> are then cleaved at <scene name='75/752344/Caspase-6_cleavage_sites_real/1'>specific sites</scene> to assume its active conformation. Zymogen activation through cleavage is largely conserved across the caspase family. However, Caspase-6 is unique in that it becomes active through self-cleavage in addition to cleavage by a separate enzymes<ref name="zincmedallinhib">PMID: 22891250 </ref>. Each monomeric unit of zymogen contains a <scene name='75/752344/Caspase-6_small_subunit_mnmr/1'>small subunit</scene> consisting of two helices, a <scene name='75/752344/Caspase-6_large_real_yeahboi/1'>large subunit</scene> consisting of three helices, a <scene name='75/752344/Caspase-6_prodomain/1'>prodomain</scene>, and a <scene name='75/752344/Caspase-6_zymogen_b-sheet/1'>beta sheet core</scene>. After cleavage at all sites, the processed post-zymogen monomers remain closely associated together through intermolecular forces as a dimer. |
| | | | |
| - | [[Image:Caspase-6 protein.jpg|100 px|left|thumb|Figure Legend]]
| + | =='''Zymogen'''== |
| | | | |
| - | [[Image:4FXO.PNG|100 px|left|thumb|This is the figure legend of the thumbnail]]
| + | In addition to a self-cleavage mechanism, Caspase-6 <scene name='75/752344/Caspase-6_zymogen_yeahboi/1'>zymogen</scene> can be activated through cleavage by Caspase-3, as well as other enzymes. This activation by cleavage is highly conserved across the caspase family, but activation through self-cleavage is uniquely recognized as the primary mechanism for Caspase-6 activation. In this self-cleavage mechanism, cleavage must occur at <scene name='75/752344/Caspase-6_cleavage_sites_real/1'>three sites</scene> in order to remove the <scene name='75/752344/Caspase-6_prodomain/1'>pro-domain</scene> located at the N-terminus and the <scene name='75/752344/Caspase-6_intersubunit_linker/1'>intersubunit linker</scene> located within the protein. These cleavages are both sequence specific and ordered, starting with cleavage of the pro-domain at <scene name='75/752344/Caspase-6_prodomain_cleavage/1'>residue 30</scene>. Removal of the intersubunit linker then occurs through cleavage at two sites, <scene name='75/752344/Caspase-6_176-179_cleavageyis/1'>DVVD179 and TEVD193</scene><ref name="RegMechStructure">PMID: 24419379 </ref>. It has been proposed that this sequence of cleavage is due to the pro-domain being more readily available to enter the active site, whose presence inhibits Caspase-6's ability to cleave the intersubunit loop and self-activate; The prodomain acts as a “suicide protector”, preventing the TEVD193 cleavage site from the active site<ref name="ActRegofCas6inND">PMID: 25340928 </ref>. After both cleavages occur, <scene name='75/752344/Active_caspase_6_dimer/1'>active Caspase-6</scene> remains in solution as a dimer. |
| - | =='''Structure'''==
| + | |
| - | ===Active Site===
| + | |
| - | In order to function as an endoprotease, Caspase-6 binds a <scene name='75/752344/Protein_ligand_real/1'>ligand</scene>, which can include neuronal proteins and tubulins [https://en.wikipedia.org/wiki/Tubulin], in its active site.[[Image:Binding grove active caspase 6.png]] This binding groove contains three critical amino acid residues necessary to perform cleavage of the peptide bonds. Together,<scene name='75/752344/His121_real/1'>His-121</scene>, <scene name='75/752344/Glu123_real/1'>Glu-123</scene>, and <scene name='75/752344/Cys163_real/1'>Cys-163</scene> form a <scene name='75/752344/Catalytic_triad_real/1'>catalytic triad</scene>[[Image:The real caspase mechanism.jpg|100 px|left|thumb|Cystine Aspartase mechanism]]. In the theorized mechanism, His-121 acts as an acid catalyst, Glu-123 acts as a base catalyst to deprotonate Cys-163, which then acts as covalent catalyst. | + | |
| | | | |
| - | ===Zinc Exosite=== | + | =='''Active State'''== |
| - | Caspase-6 function is inhibited by the binding of a <scene name='75/752344/Zinc_caspase-6/1'>Zinc</scene> ion, which binds to an <scene name='75/752344/Caspase6_allosteric_site/1'>allosteric site</scene> instead of the <scene name='75/752344/Caspase6_allostericactiv_site/1'>active site</scene>. This allosteric site is located on the outside of the protein and it is distal to the active site. The Zinc ion is bound to <scene name='75/752344/Caspase6_allosteric_site_resid/1'>three residues</scene>, Lys-36, Glu-244, and His-287, once the ion is bound to the protein it is then stabilized by a <scene name='75/752344/H20_zinc_binding_casp/1'>water molecule</scene>. The binding of Zinc at the exosite is proposed to cause a conformational change in the protein from an <scene name='75/752344/Catalytic_triad_real/1'>active state</scene> to an <scene name='75/752344/Inactive_catalytic_triad_casp/1'>inactive state</scene> that misaligns catalytic residues and inhibits activity of the enzyme. Zinc binding to the exosite is tightly regulated, because it inhibits Caspase-6's ability to inititate apoptosis. | + | [[Image:Binding grove active caspase 6.png|100 px|right|thumb|Substrate binding groove in Caspase-6. Blue - catalytic residues |
| | + | yellow - ligand |
| | + | red - generic surface]] |
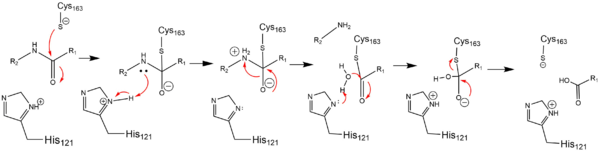
| | + | In order to function as an endopeptidase, each <scene name='75/752344/Active_caspase_6_monomer/1'>monomer</scene> of active Caspase-6 utilizes a <scene name='75/752344/Catalytic_triad_real/1'>catalytic triad</scene> composed of <scene name='75/752344/Catalytic_his-121_monomer/1'>His-121</scene>, <scene name='75/752344/Catalytic_glu-123_monomer/1'>Glu-123</scene>, and <scene name='75/752344/Catalytic_cys-163_monomer/1'>Cys-163</scene> to cleave polypeptide ligands that can include neuronal proteins and [https://en.wikipedia.org/wiki/Tubulin tubulins]<ref name="ActiveStateCrys">PMID: 21917678 </ref>. In the theorized mechanism, atoms are shown in their resting ionic states; His-121 acts as an acid catalyst, Glu-123 acts as a base catalyst to deprotonate Cys-163, which then acts as covalent catalyst. |
| | | | |
| - | =='''Activation of Caspase-6'''== | + | [[Image:Cystine Aspartase.png|600 px|active site mechanism]] |
| - | Before Caspase-6 is a functional and active dimer, the enzyme exists as a <scene name='75/752344/Caspase-6_zymogen/1'>procaspase</scene>, also known as zymogen [https://en.wikipedia.org/wiki/Zymogen]. Caspase-6 can be activated by acting as a substrate for other caspases, particularly Caspase-3, and enzymes. It becomes cleaved by these enzymes and proceeds to its <scene name='75/752344/Active_caspase_6/1'>active dimer conformation</scene>. It was observed that Caspase-6 became active without alternate enzymes present, which suggested that Caspase-6 utilizes a self-cleavage mechanism. Now, self-processing, a characteristic unique to Caspase-6, is recognized as the primary mechanism for Caspase-6 activation. The unprocessed enzyme contains a <scene name='75/752344/Caspase-6_small_subunit/1'>small</scene> and <scene name='75/752344/Caspase-6_large_real/1'>large</scene> subunit, a <scene name='75/752344/Caspase-6_pro-domain/1'>pro-domain</scene>, as well as an intersubunit linker. To become active, the intersubunit linker binds to the active site, where it is then cleaved. Other cleavages must occur as well for the enzyme to become active, specifically at TETD23 (these residues are not visible in the crystallized structure) of the pro-domain, <scene name='75/752344/Caspase-6_176-179_cleavage/1'>DVVD179</scene>, and <scene name='75/752344/Caspase-6_tevd193/1'>TEVD193</scene> amino acid sequences. Cleavage at these sites occurs in a <scene name='75/752344/Caspase-6_cleavage_sites/1'>specific sequence</scene>. First, the site within the pro-domain, TETD23, must be cleaved. This cleavage is then followed by either DVVD179 or TEVD193.
| + | =='''Zinc Inhibition'''== |
| - | Despite the sequence similarities between TETD23 and TEVD193 cleavage sites, the TETD23 cleavage site is always cleaved before TEVD193. It has been proposed that this sequence of cleavage is due to the <scene name='75/752344/Caspase-6_pr-domai-active-site/1'>structure of Caspase-6's zymogen</scene>, which allows the pro-domain to be more readily available to enter the active site. To some extent, the pro-domain inhibits Caspase-6's ability to cleave the intersubunit loop and self-activate, but this happens in a currently unknown mechanism. The result of the TETD23 cleavage site priority is that the prodomain acts as a “suicide protector”, which protects the TEVD193 cleavage site from self-cleavage[https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2966951/]. This protection is necessary when there are low levels of proteins, which must be preserved, in the tissue. The pro-domain is released after the cleavage at TETD23 and cleavage of the intersubunit links follow. then allows the two subunits to interact to form the active dimer. The intramolecular cleavage of TEVD193 is essential for the initiation caspase-6 activation without other caspases, Caspase-3 in particular, present.
| + | Caspase-6 can also assume an inactive state, which exists as a <scene name='75/752344/Casp_6_inactive_dimer/1'>dimer</scene> in its biological unit. For each <scene name='75/752344/Casp_6_inactive_monomer/1'>monomer</scene>, Caspase-6 function is primarily inhibited by the binding of a <scene name='75/752344/Casp_6_inactive_monomer_zinc/1'>zinc</scene> ion, which binds to an <scene name='75/752344/Casp_6_alosteric_site/1'>allosteric site</scene> instead of the <scene name='75/752344/Casp_6_alostericactive_site/1'>active site</scene>. This allosteric site is located on the opposite side of the protein relative to the active site. The zinc ion is bound to <scene name='75/752344/Caspase6_alloster_resid/1'>three residues</scene>, Lys-36, Glu-244, and His-287. Once the ion is bound to the protein, it is then stabilized by a <scene name='75/752344/Caspase6_alloster_h20/1'>water molecule</scene> found in the cytoplasm. The binding of zinc at the exosite is suggested to cause a conformational change in the protein from an <scene name='75/752344/Catalytic_triad_real_monomer/1'>active state</scene> to an <scene name='75/752344/Casp_6_inactive_cat_triad/1'>inactive state</scene> that misaligns catalytic residues and inhibits activity of the enzyme. It has been proposed that helices of the active dimer must rotate or move in some other way to provide these ideal interactions with zinc. This subtle shift is most likely the cause for allosteric inhibition<ref name="zincmedallinhib">PMID: 22891250 </ref>. As the helices move to bind zinc, the amino acids of the active site become misaligned. The altered positions of the amino acids no longer provide ideal interactions for incoming substrates. After zinc binds, substrates may still enter the active site, but no catalytic activity will occur. |
| | | | |
| | + | [[Image:4FXO-FINAL.jpg|300px|Caspase-6 w/ Zinc Bound]] [[Image:3s70.jpg|300px|Caspase-6 w/o Zinc Bound]] |
| | + | The first image shows the catalytic triad of Caspase-6 with zinc bound, and the second image shows the catalytic triad of caspase-6 without zinc bound. The catalytic cysteine and glutamate residues flip positions and become misaligned resulting in a loss of enzymatic function. |
| | | | |
| - | =='''Inhibition'''== | + | =='''Phosphorylation'''== |
| - | ===Zinc Inhibition===
| + | The function of Caspase-6 can be inhibited by phosphorylation of Ser-257. The exact mechanism of this reaction remains unidentified at the time of publication, but proceeds when ARK5 kinase is present. This modification can occur before and after zymogen activation. The phosphoryl group inhibits Caspase-6 through steric interference. When Ser-257 is phosphorylated, the amino acid residue interacts with <scene name='75/752344/Caspase-6_his-208/1'>Pro-201</scene>, causing a shift in the helices of Caspase-6<ref name="ActRegofCas6inND">PMID: 25340928 </ref>. This is shown in the <scene name='75/752344/Caspase-6_s257d_mutantboi/1'>S257D Caspase-6 mutant</scene>, whose mutation mimics phosphorylation<ref name="Phosregcasp6subsbindgroove">PMID: 22483120 </ref>. The shift misaligns and disrupts residues found in the active site. This conformational difference prevents the intersubunit linker from entering during zymogen activation and the self-cleaved active dimer cannot be formed. Additionally, no new substrate is able to enter the active site. |
| - | Primary inhibition of Caspase-6 occurs when a zinc ion binds to the exosite containing Lys-36, Glu-244, and His-287 of the active dimer. In addition to these residues, the zinc interacts with one water molecule from the cytoplasm. It has been proposed that helices of the active dimer must rotate or move in some other way to provide these ideal interactions with zinc. This subtle shift is most likely the cause for allosteric inhibition. As the helices move to bind zinc, the amino acids of the active site become misaligned. The altered positions of the amino acids no longer provide ideal interactions for incoming substrates. After zinc binds, no new substrates enter the active site. Thus, Caspase-6 is effectively inhibited.
| + | |
| | | | |
| - | ===Phosphorylation=== | + | =='''Medical Relevance'''== |
| - | The function of Caspase-6 can be inhibited by phosphorylation of Ser-257. The exact mechanism of this reaction remains unidentified at the time of publication, but proceeds when ARK5 kinase is present. This modification can occur before and after zymogen activation or auto-processing. The phosphoryl group inhibits Caspase-6 through steric interference. When Ser-257 is phosphorylated, the amino acid residue interacts with Pro-201, causing a shift in the helices of Caspase-6. This is shown in the <scene name='75/752344/Caspase-6_s257d_mutant/1'>S257D Caspase-6 mutant</scene> mutant, whose mutation mimics phosphorylation. The shift misaligns and disrupts residues found in the active site. This conformational difference prevents the inter-subunit loop from entering during zymogen activation and the self-cleaved active dimer cannot be formed. Additionally, no new substrate is able to enter the active site.
| + | |
| - | | + | |
| - | | + | |
| - | =='''Function'''==
| + | |
| | ===Caspase-6 involvement in Alzheimer's Disease=== | | ===Caspase-6 involvement in Alzheimer's Disease=== |
| - | Caspase-6 activity is associated with the formation of lesions within the Alzheimer's Disease (AD) and they can become present very early on during the disease's progression. Proapoptotic protein p53 is present at increased levels within AD brains, which seems to directly increase the transcription of Caspase-6. Treatments of Alzheimer's include targeting active Caspase-6 proteins because staining has found active Caspase-6 within the hippocampus and cortex of the Brain within in mild, moderate, and severe cases of AD, which indicates that Caspase-6 plays a predominate role in the pathophysiology of Alzheimer's. There has been research conducted that shows activation of Caspase-6 in AD could cause disruption of the cytoskeleton network of neurons, which then causes handicapped synaptic plasticity. | + | Caspase-6 is known to be involved in many neurodegenerative diseases, one of which is Alzheimer's disease (AD). Caspase-6 activity is associated with the formation of lesions within the [http://www.alz.org/ Alzheimer's Disease].Lesions can be found in early stages of AD. A proapoptotic protein, p53, is present at increased levels within AD brains, which seems to directly increase the transcription of Caspase-6, which indirectly influences apoptosis of neurons. Future treatments of AD include selective inhibition of active Caspase-6 proteins; staining has found active Caspase-6 within the hippocampus and cortex of the brain within a varying severity of AD cases. This suggests that Caspase-6 plays a predominate role in the pathophysiology of AD. There has been research conducted that shows activation of Caspase-6 in AD could cause disruption of the cytoskeleton network of neurons and lead to neuronal apoptosis<ref name="ActRegofCas6inND">PMID: 25340928 </ref>. |
| - | | + | |
| - | | + | |
| - | | + | |
| - | | + | |
| - | ==Luke's free space==
| + | |
| - | If <scene name='75/752344/Serine_257_highlighted/1'>Serine 257</scene> is <scene name='pdbligand=PO4:PHOSPHATE+ION'>Phospohrylated</scene> , the activity of this protein is inhibited.
| + | |
| - | [[Image:Caspase 6 sequence image key.JPG]]
| + | |
| - | [[Image:Caspase 6 sequence image.JPG]]
| + | |
| - | If <scene name='pdbligand=ZN:ZINC+ION'>Zinc</scene> binds to the protein, the activity of the active site is inhibited.
| + | |
| - | | + | |
| - | Inactive state of caspase 6:
| + | |
| - | <scene name='75/752344/Uncleaved_caspase_6/1'>inactive caspase</scene>
| + | |
| - | | + | |
| - | == Relevance ==
| + | |
| - | | + | |
| - | </StructureSection>
| + | |
| - | == References ==
| + | |
| - | <references/>
| + | |
| - | Wang, Xiao-Jun, Qin Cao, Yan Zhang, and Xiao-Dong Su. "Activation and Regulation of Caspase-6 and Its Role in Neurodegenerative Diseases." Annual Review of Pharmacology and Toxicology 55.1 (2015): 553-72. Web.
| + | |
| - | | + | |
| - | Wang XJ, Cao Q, Liu X, Wang KT, Mi W, et al. 2010. Crystal structures of human caspase 6 reveal a new mechanism for intramolecular cleavage self-activation. EMBO Rep. 11: 841–47
| + | |
| - | (self cleavage article)
| + | |
| | | | |
| - | http://www.rcsb.org/pdb/explore/explore.do?structureId=2WDP (this is the non-self cleaved protien)
| + | =='''References'''== |
| | + | {{reflist}} |
| Caspase-6 is an endopeptidase involved in apoptosis. In terms of its catalytic function, it is a part of the cysteine-aspartate family. Before Caspase-6 becomes functional, the enzyme exists as a , also known as a zymogen. This zymogen exists as a , whose are then cleaved at to assume its active conformation. Zymogen activation through cleavage is largely conserved across the caspase family. However, Caspase-6 is unique in that it becomes active through self-cleavage in addition to cleavage by a separate enzymes[1]. Each monomeric unit of zymogen contains a consisting of two helices, a consisting of three helices, a , and a . After cleavage at all sites, the processed post-zymogen monomers remain closely associated together through intermolecular forces as a dimer.
Zymogen
In addition to a self-cleavage mechanism, Caspase-6 can be activated through cleavage by Caspase-3, as well as other enzymes. This activation by cleavage is highly conserved across the caspase family, but activation through self-cleavage is uniquely recognized as the primary mechanism for Caspase-6 activation. In this self-cleavage mechanism, cleavage must occur at in order to remove the located at the N-terminus and the located within the protein. These cleavages are both sequence specific and ordered, starting with cleavage of the pro-domain at . Removal of the intersubunit linker then occurs through cleavage at two sites, [2]. It has been proposed that this sequence of cleavage is due to the pro-domain being more readily available to enter the active site, whose presence inhibits Caspase-6's ability to cleave the intersubunit loop and self-activate; The prodomain acts as a “suicide protector”, preventing the TEVD193 cleavage site from the active site[3]. After both cleavages occur, remains in solution as a dimer.
Active State
 Substrate binding groove in Caspase-6. Blue - catalytic residues yellow - ligand red - generic surface In order to function as an endopeptidase, each of active Caspase-6 utilizes a composed of , , and to cleave polypeptide ligands that can include neuronal proteins and tubulins[4]. In the theorized mechanism, atoms are shown in their resting ionic states; His-121 acts as an acid catalyst, Glu-123 acts as a base catalyst to deprotonate Cys-163, which then acts as covalent catalyst.
Zinc Inhibition
Caspase-6 can also assume an inactive state, which exists as a in its biological unit. For each , Caspase-6 function is primarily inhibited by the binding of a ion, which binds to an instead of the . This allosteric site is located on the opposite side of the protein relative to the active site. The zinc ion is bound to , Lys-36, Glu-244, and His-287. Once the ion is bound to the protein, it is then stabilized by a found in the cytoplasm. The binding of zinc at the exosite is suggested to cause a conformational change in the protein from an to an that misaligns catalytic residues and inhibits activity of the enzyme. It has been proposed that helices of the active dimer must rotate or move in some other way to provide these ideal interactions with zinc. This subtle shift is most likely the cause for allosteric inhibition[1]. As the helices move to bind zinc, the amino acids of the active site become misaligned. The altered positions of the amino acids no longer provide ideal interactions for incoming substrates. After zinc binds, substrates may still enter the active site, but no catalytic activity will occur.
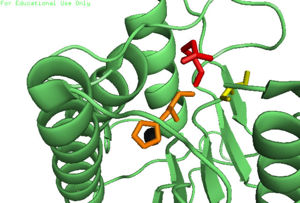 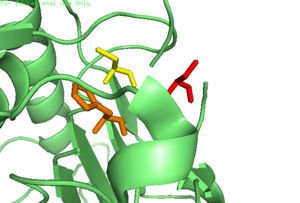
The first image shows the catalytic triad of Caspase-6 with zinc bound, and the second image shows the catalytic triad of caspase-6 without zinc bound. The catalytic cysteine and glutamate residues flip positions and become misaligned resulting in a loss of enzymatic function.
Phosphorylation
The function of Caspase-6 can be inhibited by phosphorylation of Ser-257. The exact mechanism of this reaction remains unidentified at the time of publication, but proceeds when ARK5 kinase is present. This modification can occur before and after zymogen activation. The phosphoryl group inhibits Caspase-6 through steric interference. When Ser-257 is phosphorylated, the amino acid residue interacts with , causing a shift in the helices of Caspase-6[3]. This is shown in the , whose mutation mimics phosphorylation[5]. The shift misaligns and disrupts residues found in the active site. This conformational difference prevents the intersubunit linker from entering during zymogen activation and the self-cleaved active dimer cannot be formed. Additionally, no new substrate is able to enter the active site.
Medical Relevance
Caspase-6 involvement in Alzheimer's Disease
Caspase-6 is known to be involved in many neurodegenerative diseases, one of which is Alzheimer's disease (AD). Caspase-6 activity is associated with the formation of lesions within the Alzheimer's Disease.Lesions can be found in early stages of AD. A proapoptotic protein, p53, is present at increased levels within AD brains, which seems to directly increase the transcription of Caspase-6, which indirectly influences apoptosis of neurons. Future treatments of AD include selective inhibition of active Caspase-6 proteins; staining has found active Caspase-6 within the hippocampus and cortex of the brain within a varying severity of AD cases. This suggests that Caspase-6 plays a predominate role in the pathophysiology of AD. There has been research conducted that shows activation of Caspase-6 in AD could cause disruption of the cytoskeleton network of neurons and lead to neuronal apoptosis[3].
References
- ↑ 1.0 1.1 Velazquez-Delgado EM, Hardy JA. Zinc-Mediated Allosteric Inhibition of Caspase-6. J Biol Chem. 2012 Aug 13. PMID:22891250 doi:http://dx.doi.org/10.1074/jbc.M112.397752
- ↑ Cao Q, Wang XJ, Li LF, Su XD. The regulatory mechanism of the caspase 6 pro-domain revealed by crystal structure and biochemical assays. Acta Crystallogr D Biol Crystallogr. 2014 Jan;70(Pt 1):58-67. doi:, 10.1107/S1399004713024218. Epub 2013 Dec 24. PMID:24419379 doi:http://dx.doi.org/10.1107/S1399004713024218
- ↑ 3.0 3.1 3.2 Wang XJ, Cao Q, Zhang Y, Su XD. Activation and regulation of caspase-6 and its role in neurodegenerative diseases. Annu Rev Pharmacol Toxicol. 2015;55:553-72. doi:, 10.1146/annurev-pharmtox-010814-124414. Epub 2014 Oct 17. PMID:25340928 doi:http://dx.doi.org/10.1146/annurev-pharmtox-010814-124414
- ↑ Betz BL, Roh MH, Weigelin HC, Placido JB, Schmidt LA, Farmen S, Arenberg DA, Kalemkerian GP, Knoepp SM. The application of molecular diagnostic studies interrogating EGFR and KRAS mutations to stained cytologic smears of lung carcinoma. Am J Clin Pathol. 2011 Oct;136(4):564-71. doi: 10.1309/AJCP84TUTQOSUONG. PMID:21917678 doi:http://dx.doi.org/10.1309/AJCP84TUTQOSUONG
- ↑ Velazquez-Delgado EM, Hardy JA. Phosphorylation regulates assembly of the caspase-6 substrate-binding groove. Structure. 2012 Apr 4;20(4):742-51. Epub 2012 Apr 3. PMID:22483120 doi:10.1016/j.str.2012.02.003
|




