Aspartate Aminotransferase
From Proteopedia
(Difference between revisions)
| (9 intermediate revisions not shown.) | |||
| Line 1: | Line 1: | ||
<StructureSection load='1asn' size='400' side='right' scene='' caption='E. coli aspartate aminotransferase complex with PLP and sulfate (PDB code [[1asn]])'> | <StructureSection load='1asn' size='400' side='right' scene='' caption='E. coli aspartate aminotransferase complex with PLP and sulfate (PDB code [[1asn]])'> | ||
| - | + | '''Aspartate Aminotransferase''' | |
by Luke Spooner | by Luke Spooner | ||
| - | '''Aspartate Aminotransferase''' (AAT), also known as Glutamic aspartic transaminase, glutamic oxaloacetic transaminase, and transaminase A | + | __TOC__ |
| + | |||
| + | ==Function== | ||
| + | |||
| + | '''Aspartate Aminotransferase''' (AAT), also known as '''Glutamic aspartic transaminase''', '''glutamic oxaloacetic transaminase''', '''prephenate aminotransferase''' and '''transaminase A''' is an enzyme that is a member of the class-I pyridoxal-phosphate-dependent aminotransferase family <ref name ="AST family and name">PMID:20977429</ref>. It is coded by the gene GOT1<ref name ="AST gene">PMID:4193185</ref>. It is a homodimer that is 413 amino acids long and serves a critical role in amino acid and carbohydrate metabolism, ureogenesis, and the transfer of reducing equivalents into the mitochondria and chloroplast<ref name ="AST ROLES AND STRUCTURE">PMID:10708649</ref>. Within prokaryote cells it is exclusively found in the cytosol, but in eukaryotic cells there are cytosol, mitochondrial, and chloroplast isozymes<ref name ="AST family and name"/><ref name ="AST Structure"/>. | ||
| + | *'''Bifunctional aspartate aminotransferase''' (BAAT) is found in plants. It displays aspartate and prephanate aminotransferase activity<ref >PMID:24902885</ref>. | ||
In the human body it is produced in the brain, skeletal muscles, liver, pancreas, red blood cells, and kidneys <ref name ="AST ORGANS">PMID:2569674</ref><ref name ="Liver damage"/>. The wide range of tissues in which it is made, separates it from the similar enzyme alanine transaminase (ALT) which is found primarily in the liver<ref name ="Liver damage">PMID:10831269</ref>. The level of AAT in the body can be used as a marker for tissue disease or damage<ref name ="Liver damage"/>. As well, AAT and ALT levels can be compared to pinpoint whether tissue damage is primarily found within the liver<ref name ="Liver damage">PMID:12546613</ref>. | In the human body it is produced in the brain, skeletal muscles, liver, pancreas, red blood cells, and kidneys <ref name ="AST ORGANS">PMID:2569674</ref><ref name ="Liver damage"/>. The wide range of tissues in which it is made, separates it from the similar enzyme alanine transaminase (ALT) which is found primarily in the liver<ref name ="Liver damage">PMID:10831269</ref>. The level of AAT in the body can be used as a marker for tissue disease or damage<ref name ="Liver damage"/>. As well, AAT and ALT levels can be compared to pinpoint whether tissue damage is primarily found within the liver<ref name ="Liver damage">PMID:12546613</ref>. | ||
__TOC__ | __TOC__ | ||
=='''Structure'''== | =='''Structure'''== | ||
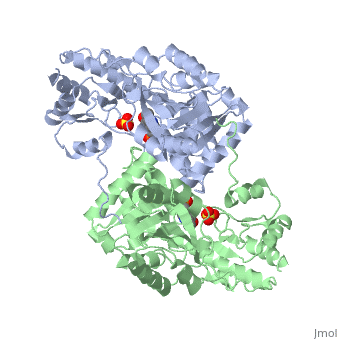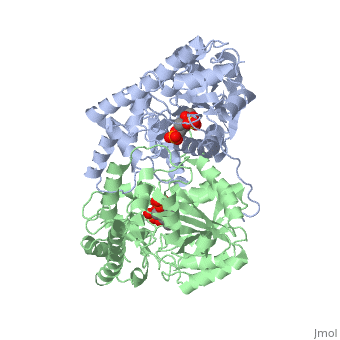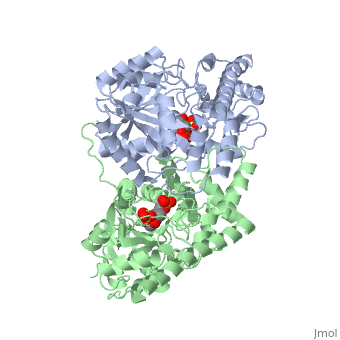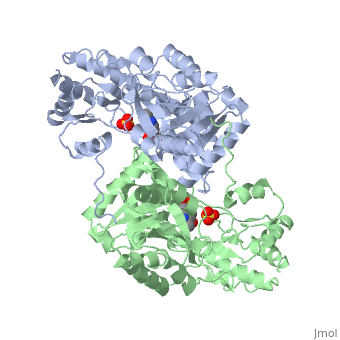
| - | AST is a homodimer that contains <scene name=' | + | <scene name='49/490061/Cv/1'>AST is a homodimer</scene> that contains <scene name='49/490061/Cv/3'>16 α-helices and a β-sheet formed from 7 parallel and antiparallel strands</scene> <ref name ="AST Structure"/> ({{Template:ColorKey_Helix}}, |
| + | {{Template:ColorKey_Strand}}, {{Template:ColorKey_Loop}}, {{Template:ColorKey_Turn}}). Asymmetric unit of Aspartate aminotransferase, with highlighted small and large domain and PLP cofactor ([[1b4x]]). Each subunit contains an equivalent active site<ref name ="AST Structure">PMID:2121725</ref>. The subunits connect at two sites: between their large domains and between the N-terminal residues and the large domain on the other subunit. This structure of AST varies minutely among organisms ranging from ''E. coli'' to humans<ref name ="AST Structure"/><ref name ="AST ROLES AND STRUCTURE"/>. As well, the structure of the active site is highly conserved with a sequence homology of 25%<ref name ="AST Structure"/>. | ||
Each subunit of the homodimer is further divided into a small and large domain<ref name ="AST Structure"/>. The <scene name='Sandbox_Reserved_346/Small_subunit_2/1'>small domain</scene> is comprised of the amino acids from the N-terminus to Pro 48 residue and from Met 326 to the C-terminus<ref name ="AST Structure"/>. The remaining amino acids make up the <scene name='Sandbox_Reserved_346/Large_subunit/1'>large domain</scene>, and the <scene name='Sandbox_Reserved_346/Whole_subunit_2/1'>two domains</scene> are connected by a long α-helix consisting of 32 amino acids<ref name ="AST Structure"/>. | Each subunit of the homodimer is further divided into a small and large domain<ref name ="AST Structure"/>. The <scene name='Sandbox_Reserved_346/Small_subunit_2/1'>small domain</scene> is comprised of the amino acids from the N-terminus to Pro 48 residue and from Met 326 to the C-terminus<ref name ="AST Structure"/>. The remaining amino acids make up the <scene name='Sandbox_Reserved_346/Large_subunit/1'>large domain</scene>, and the <scene name='Sandbox_Reserved_346/Whole_subunit_2/1'>two domains</scene> are connected by a long α-helix consisting of 32 amino acids<ref name ="AST Structure"/>. | ||
| Line 26: | Line 32: | ||
=='''Clinical Applications'''== | =='''Clinical Applications'''== | ||
The levels of AAT in the body are indicative of tissue damage and disease<ref name ="TISSUE DAMAGE">PMID:8432855</ref>. Normally AAT is found in minimal amounts within the blood, however when the organs mentioned above are damaged, AAT is released into the blood<ref name ="TISSUE DAMAGE"/>. The amount released is proportional to the level of damage sustained<ref name ="TISSUE DAMAGE"/>. AAT levels have been shown to rise substantially within 6 hours of the initial tissue degradation and can stay elevated for up to 4 days<ref name ="TISSUE DAMAGE"/>. AAT levels when compared with the levels of other enzymes can be used by physicians to determine where in the body the damage has taken place<ref name ="Liver damage"/>. Comparisons with ALT have proven particularly useful in identifying liver damage such as cirrhosis and hepatitis<ref name ="Liver damage"/>. Under normal condition, AAT levels within men are 6-34 IU/L and for women it is 8-40 IU/L<ref name ="TISSUE DAMAGE"/>. | The levels of AAT in the body are indicative of tissue damage and disease<ref name ="TISSUE DAMAGE">PMID:8432855</ref>. Normally AAT is found in minimal amounts within the blood, however when the organs mentioned above are damaged, AAT is released into the blood<ref name ="TISSUE DAMAGE"/>. The amount released is proportional to the level of damage sustained<ref name ="TISSUE DAMAGE"/>. AAT levels have been shown to rise substantially within 6 hours of the initial tissue degradation and can stay elevated for up to 4 days<ref name ="TISSUE DAMAGE"/>. AAT levels when compared with the levels of other enzymes can be used by physicians to determine where in the body the damage has taken place<ref name ="Liver damage"/>. Comparisons with ALT have proven particularly useful in identifying liver damage such as cirrhosis and hepatitis<ref name ="Liver damage"/>. Under normal condition, AAT levels within men are 6-34 IU/L and for women it is 8-40 IU/L<ref name ="TISSUE DAMAGE"/>. | ||
| - | + | ||
==3D structures of aspartate aminotransferase== | ==3D structures of aspartate aminotransferase== | ||
| + | [[Aspartate aminotransferase 3D structures]] | ||
| - | + | </StructureSection> | |
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
| - | + | ||
Current revision
| |||||||||||

References
- ↑ 1.0 1.1 Han Q, Robinson H, Cai T, Tagle DA, Li J. Biochemical and structural characterization of mouse mitochondrial aspartate aminotransferase, a newly identified kynurenine aminotransferase-IV. Biosci Rep. 2010 Oct 26. PMID:20977429 doi:10.1042/BSR20100117
- ↑ DeLorenzo RJ, Ruddle FH. Glutamate oxalate transaminase (GOT) genetics in Mus musculus: linkage, polymorphism, and phenotypes of the Got-2 and Got-1 loci. Biochem Genet. 1970 Apr;4(2):259-73. PMID:4193185
- ↑ 3.0 3.1 3.2 3.3 3.4 3.5 Jeffery CJ, Gloss LM, Petsko GA, Ringe D. The role of residues outside the active site: structural basis for function of C191 mutants of Escherichia coli aspartate aminotransferase. Protein Eng. 2000 Feb;13(2):105-12. PMID:10708649
- ↑ 4.00 4.01 4.02 4.03 4.04 4.05 4.06 4.07 4.08 4.09 4.10 4.11 4.12 4.13 4.14 4.15 4.16 4.17 4.18 4.19 Kamitori S, Okamoto A, Hirotsu K, Higuchi T, Kuramitsu S, Kagamiyama H, Matsuura Y, Katsube Y. Three-dimensional structures of aspartate aminotransferase from Escherichia coli and its mutant enzyme at 2.5 A resolution. J Biochem. 1990 Aug;108(2):175-84. PMID:2121725
- ↑ de la Torre F, Cañas RA, Pascual MB, Avila C, Cánovas FM. Plastidic aspartate aminotransferases and the biosynthesis of essential amino acids in plants. J Exp Bot. 2014 Oct;65(19):5527-34. PMID:24902885 doi:10.1093/jxb/eru240
- ↑ Palaiologos G, Hertz L, Schousboe A. Role of aspartate aminotransferase and mitochondrial dicarboxylate transport for release of endogenously and exogenously supplied neurotransmitter in glutamatergic neurons. Neurochem Res. 1989 Apr;14(4):359-66. PMID:2569674
- ↑ 7.0 7.1 7.2 7.3 7.4 7.5 Tran A, Longo F, Ouzan D, Bianchi D, Pradier C, Saint-Paul MC, Sattonnet C, Laffont C, Dantin S, Piche T, Benzaken S, Rampal P. Effects of 1-year interferon-alpha 2a treatment in patients with chronic hepatitis C and persistently normal transaminase activity. Scand J Gastroenterol. 2000 Apr;35(4):433-7. PMID:10831269
- ↑ 8.0 8.1 8.2 8.3 8.4 Martinez-Carrion M, Tiemeier DC, Peterson DL. Conformational properties of the isoenzymes of aspartate transaminase and the enzyme-substrate complexes. Biochemistry. 1970 Jun 23;9(13):2574-82. PMID:5450225
- ↑ 9.0 9.1 Tretter L, Adam-Vizi V. Inhibition of Krebs cycle enzymes by hydrogen peroxide: A key role of [alpha]-ketoglutarate dehydrogenase in limiting NADH production under oxidative stress. J Neurosci. 2000 Dec 15;20(24):8972-9. PMID:11124972
- ↑ 10.0 10.1 Tretter L, Adam-Vizi V. Inhibition of Krebs cycle enzymes by hydrogen peroxide: A key role of [alpha]-ketoglutarate dehydrogenase in limiting NADH production under oxidative stress. J Neurosci. 2000 Dec 15;20(24):8972-9. PMID:11124972
- ↑ 11.0 11.1 Jungas RL, Halperin ML, Brosnan JT. Quantitative analysis of amino acid oxidation and related gluconeogenesis in humans. Physiol Rev. 1992 Apr;72(2):419-48. PMID:1557428
- ↑ 12.0 12.1 Gibbs ME, Hertz L. Importance of glutamate-generating metabolic pathways for memory consolidation in chicks. J Neurosci Res. 2005 Jul 15;81(2):293-300. PMID:15929064 doi:10.1002/jnr.20548
- ↑ 13.0 13.1 13.2 13.3 13.4 Gonzalez-Flecha B, Cutrin JC, Boveris A. Time course and mechanism of oxidative stress and tissue damage in rat liver subjected to in vivo ischemia-reperfusion. J Clin Invest. 1993 Feb;91(2):456-64. PMID:8432855 doi:http://dx.doi.org/10.1172/JCI116223
