HMG-CoA Reductase
From Proteopedia
| Line 12: | Line 12: | ||
==Structure== | ==Structure== | ||
| - | < | + | <StructureSection load='1dq8' size='500' side='right' caption='HMG-CoA_Reductase/1dq8_starting_scene/1'> |
| - | < | + | ===General Structure=== |
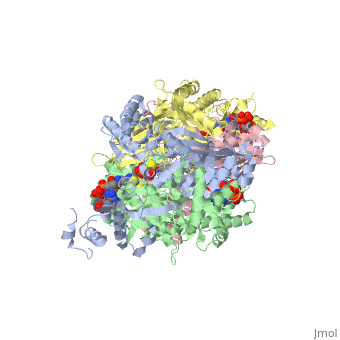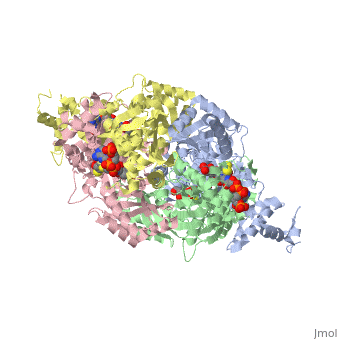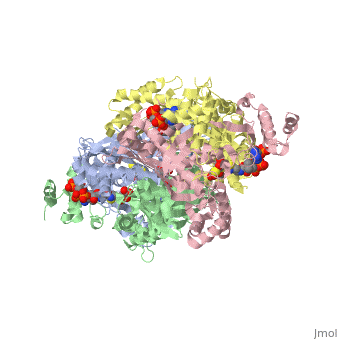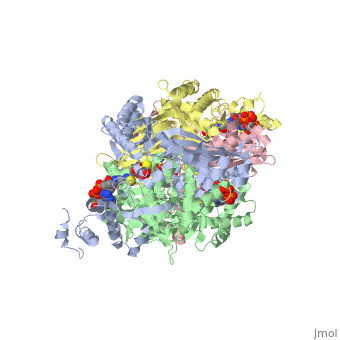
| - | < | + | There are two distinct classes of HMGRs, class I, which is only found in eukaryotes and are membrane bound and class II, which is found in prokaryotes and are soluble. <ref>PMID:11349148</ref> HMGR contains 8 transmembrane domains that have yet to be successfully crystallized, which anchor the protein to the membrane of the endoplasmic reticulum. <ref name="Roitelman"/> The catalytic portion of human HMGR forms a tetramer, with the individual monomers winding around each other. <ref name="Roitelman">PMID:1374417</ref> Within the tetramer, the monomers are arranged into <scene name='HMG-CoA_Reductase/1dq8_2_dimers/3'>two dimers</scene>, each of which contains <scene name='HMG-CoA_Reductase/1dq8_2_active_sites/2'>two active sites </scene>which are formed by residues form both monomers. Each monomer contains <scene name='HMG-CoA_Reductase/1dq8_star3_domains/2'>three domains </scene>, the <scene name='HMG-CoA_Reductase/1dq8_n_domain/2'>N-domain</scene>, the <scene name='HMG-CoA_Reductase/1dq8_l_domain/1'>L-Domain</scene>, and the <scene name='HMG-CoA_Reductase/1dq8_s_domain/1'>S-Domain</scene>. The L-domain is unique to HMGRs while the S-domain, which forms the binding site for NADP, resembles that of [[ferredoxin]]. The S and L domains are connected by a <scene name='HMG-CoA_Reductase/1dq8_cis_loop/6'>“cis-loop”</scene> which is essential for the HMG-binding site. <ref name="Roitelman"/> Salt bridges between residues R641 and E782 as well as <scene name='HMG-CoA_Reductase/1dq8_cis_loop/4'>hydrogen bonds</scene> between E700 and E700 on neighboring monomers compliment the largely hydrophobic dimer-dimer interface. <ref name="Roitelman"/> |
| - | < | + | <br /> |
| - | </ | + | |
| - | < | + | |
| - | < | + | ===Substrate Binding & Catalytic Mechanism=== |
| - | + | [[Image: Reactin_scheme.PNG|200px|left|thumb| Chemical Reaction Catalyzed by HMGR]] | |
| - | </ | + | The HMG-CoA and NADPH molecules make numerous contacts with the <scene name='HMG-CoA_Reductase/Ls_domains/3'>L and S domains</scene> in forming the four active sites. The CoA is located in a <scene name='HMG-CoA_Reductase/1dqa_nadp_and_coa/2'>near the enzyme surface</scene>, with the pantothenic acid moiety extending into the interior of the protein. <scene name='HMG-CoA_Reductase/1dqa_tyr_491/2'>Tyrosine 479 forms a hydrophobic lid</scene> over the CoA adenine base, shielding the extended binding pocket from solution. The NADPH binding site is formed primarily by the S-domain with <scene name='HMG-CoA_Reductase/1dqa_loop/2'>a loop region</scene> playing a critical role in binding. <ref name="Roitelman"/> |
| - | </ | + | |
| - | </ | + | The HMG binding pocket is the site of catalysis in HMGR. <scene name='HMG-CoA_Reductase/1dqa_cis_loop2/2'> The“cis-loop” that bends over the top of HMG </scene> is a critical structural element of this binding site. Residues <scene name='HMG-CoA_Reductase/1dqa_e_and_d/2'>E559 and D767</scene> and are positioned in the active site as is <scene name='HMG-CoA_Reductase/1dqa_k691/2'>K691 which is only 2.7 angstroms from the HMG O2 carbonyl oxygen</scene>. It is this K691 that likely stabilizes the negatively charged oxygen of the first mevaldyl-CoA intermediate. <ref name="Roitelman"/> The mevaldyl CoA intermediate is subsequently converted to Mavaldehyde with added stabilization from <scene name='HMG-CoA_Reductase/1dqa_h866/2'>H866, which is within hydrogen bonding distance of the thiol group</scene>. It is then believed that the close proximity of <scene name='HMG-CoA_Reductase/1dqa_e_and_d/2'>E559 and D767</scene> increases the pKA of E559, allowing it to be a proton donor for the reduction of mevaldehyde into mevalonate. <ref name="Roitelman"/> |
| - | </ | + | <br /> |
| + | |||
| + | ==Regulation of HMG-CoA Reductase== | ||
| + | |||
| + | As stated before, HMGR is among the most highly regulated enzymes in the human body. <ref name="Meigs">PMID:8626470</ref> Regulation of HMG-CoA reductase occurs at 4 different levels of feedback regulation. First by regulation of transcription of the reductase gene, which is activated by sterol regulatory element binding protein, a protein that binds to the promoter of the HMGR gene when cholesterol levels fall. The second level of regulation is at the translation of the HMGR mRNA, which is inhibited by Farnesol, a derivative of the mevalonate pathway. <ref name="Meigs"/> | ||
| + | |||
| + | The third level of HMGR regulation involves the degradation of intact reductase. As the level of sterols increases, HMGR reductase becomes more susceptible to ER-associated degradation. Helices 2-6 of the HMGR transmembrane domain, called the sterol-sensing domain, sense the increased levels of sterols. Degradation of HMGR is initiated when a membrane-bound cysteine protease cleaves residues within the transmembrane region of HMGR. Additionally, as sterol levels increase, the helices can expose Lysine 248 which can be ubiquitinated and subsequently trigger proteolytic degradation. <ref name="Roitelman"/> <ref>PMID:16168377</ref> | ||
| + | |||
| + | A final level of HMGR regulation is achieved by inhibition via <scene name='HMG-CoA_Reductase/1dqa_phosphorylation_871/1'>phosphorylation of Serine 872</scene>. HMGR is phosphorylated and inactivated by an AMP-activated protein kinase, when the energy charge of the cell is low and AMP concentrations are high. Since Serine 872 is in the vicinity of the pohsphate of NADP, phosphorylation likely results in a decreased affinity for NADPH, halting the formation of Mevaldyl-CoA from HMG-CoA. | ||
| + | <ref>PMID:1967820</ref><br /> | ||
| + | </StructureSection> | ||
==Medical Implications== | ==Medical Implications== | ||
Revision as of 06:04, 2 November 2010

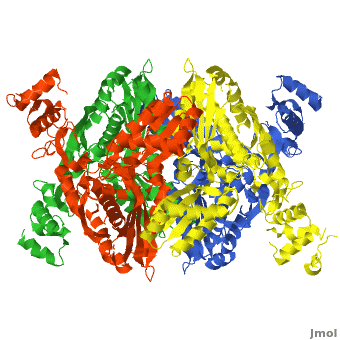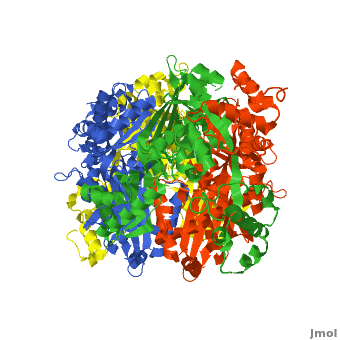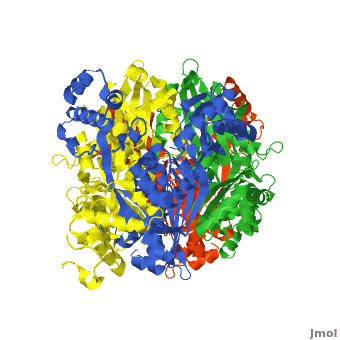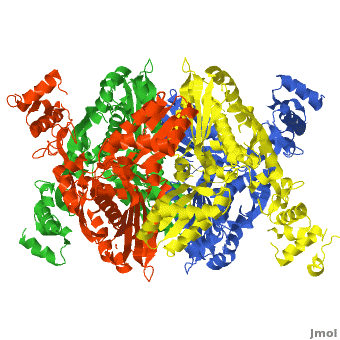

HMG-CoA Reductase (or 3-hydroxy-3-methyl-glutaryl-CoA reductasese or HMGR) is the rate-controlling enzyme of the mevalonate pathway, responsible for cholesterol and other isoprenoid biosynthesis. HMGR is a polytopic, transmembrane protein, containing 8 domains, that is anchored in the membrane of the endoplasmic reticulum.[1] It is the major target of the Statins, a cholesterol lowering drug class and the best selling pharmaceutical drugs in the world.

Contents |
Biological Role
HMGR is among the most highly regulated enzymes in the human body. It catalyzes the formation of mevalonic acid, the committed step in the biosynthesis of sterols, most notably cholesterol. This reaction can be seen below where HMG-CoA is reduced by NADPH. Despite the poor reputation cholesterol has in the media, it is a critical component of cellular membranes as it is required to establish proper membrane permeability and fluidity. The mevalonate pathway is also responsible for synthesis of the oxygen transporting heme found in red blood cells. [2]
Structure
| |||||||||||

Medical Implications
| |||||||||||

Additional 3D Structures of HMG-CoA Reductase
HGMCR
1r7i - PmHMGCR catalytic domain - Pseudomonas mevalonii
HGMCR+statins
3cct, 3ccw, 3ccz, 3cd0, 3cd5, 3cd7, 3cda, 3cdb, 2r4f, 3bgl, 2q1l, 2q6b, 2q6c, 1hw8, 1hw9, 1hwi, 1hwj, 1hwk, 1hwl – HMG-CoA Reductase Catalytic Domain + Statins
1t02 - PmHMGCR catalytic domain + statin derivatives
HGMCR+cofactors
1r31 – PmHMGCR catalytic domain +CoA+mevalonate
1qax - PmHMGCR catalytic domain +HMG+CoA+NAD+
1qay - PmHMGCR catalytic domain +mevalonate+NAD+
1dq8 - hHMGCR catalytic domain (mutant) +CoA+HMG
1dq9 - hHMGCR catalytic domain (mutant)+HMG-CoA
1dqa - hHMGCR catalytic domain (mutant)+HMG+CoA+NADP+
Additional Resources
- See: Pharmaceutical Drug Targets For Additional Information about Drug Targets for Related Diseases
- See: Metabolic Disorders For Additional Information.
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 Roitelman J, Olender EH, Bar-Nun S, Dunn WA Jr, Simoni RD. Immunological evidence for eight spans in the membrane domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase: implications for enzyme degradation in the endoplasmic reticulum. J Cell Biol. 1992 Jun;117(5):959-73. PMID:1374417
- ↑ 2.0 2.1 2.2 Meigs TE, Roseman DS, Simoni RD. Regulation of 3-hydroxy-3-methylglutaryl-coenzyme A reductase degradation by the nonsterol mevalonate metabolite farnesol in vivo. J Biol Chem. 1996 Apr 5;271(14):7916-22. PMID:8626470
- ↑ Istvan ES, Deisenhofer J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science. 2001 May 11;292(5519):1160-4. PMID:11349148 doi:10.1126/science.1059344
- ↑ Song BL, Sever N, DeBose-Boyd RA. Gp78, a membrane-anchored ubiquitin ligase, associates with Insig-1 and couples sterol-regulated ubiquitination to degradation of HMG CoA reductase. Mol Cell. 2005 Sep 16;19(6):829-40. PMID:16168377 doi:10.1016/j.molcel.2005.08.009
- ↑ Goldstein JL, Brown MS. Regulation of the mevalonate pathway. Nature. 1990 Feb 1;343(6257):425-30. PMID:1967820 doi:http://dx.doi.org/10.1038/343425a0
- ↑ www.nhlbi.nih.gov/health/.../Diseases/.../CAD_WhatIs.html
- ↑ Endo A, Kuroda M, Tanzawa K. Competitive inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase by ML-236A and ML-236B fungal metabolites, having hypocholesterolemic activity. FEBS Lett. 1976 Dec 31;72(2):323-6. PMID:16386050
- ↑ http://www.drugs.com/top200.html
- ↑ http://www.medicalnewstoday.com/articles/25046.php
- ↑ Istvan ES, Deisenhofer J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science. 2001 May 11;292(5519):1160-4. PMID:11349148 doi:10.1126/science.1059344
- ↑ Corsini A, Maggi FM, Catapano AL. Pharmacology of competitive inhibitors of HMG-CoA reductase. Pharmacol Res. 1995 Jan;31(1):9-27. PMID:7784310
Proteopedia Page Contributors and Editors (what is this?)
David Canner, Michal Harel, Alexander Berchansky, Eran Hodis, Angel Herraez, Joel L. Sussman
