Ionotropic Glutamate Receptors
From Proteopedia
(Difference between revisions)
| Line 4: | Line 4: | ||
[[Autism Spectrum Disorders]] (ASDs) are developmental disorders. During development, glutamate regulates neuronal growth and synaptogenesis, effectively dictating the underlying cytoarchitecture of the brain.<ref name="Purcel"/> Significant research into ASDs has been devoted to understanding how glutamate receptors function and how their disruption might lead to disorders. IGluRs are concentrated in regions of the brain that have been implicated in ASDs including the cerebellum and hippocampus, areas responsible for motor control, spatial navigation and memory. Studies have revealed that glutamate receptor proteins are increased in the cerebellum of autistic individuals by nearly 250% and specific small nucleotide polymorphisms in IGluRs have been identified which correlate with the ASDs. Further, many people with autism have clearly visible disturbances in the anatomy of the inferior olive (IO), a small part of the brain responsible for movement coordination and maintenance of an underlying 12 Hz brain rhythm through careful regulation of glutamate signaling.<ref>PMID: 15749250</ref> A well-known mutation in glutamate receptors in the “Lurcher” mouse model has revealed that constitutively leaky glutamate receptor ion channel result in IO neuron degeneration and loss of purkinje cells, with the mice exhibiting Autism-like characteristics.<ref>PMID: 9285588</ref> Such relationships between overly active glutamate receptors leading to increased excitation/inhibition ratios and autism have led some to propose using glutamate receptor inhibitors as a means of pharmaceutical intervention for improving those with autistic symptoms.<ref>PMID: 14606691</ref> Many pharmacological agents that reduce neural excitation, such as benzodiazapines and anticonvulsants, are thought to potentially have therapeutic value in treating autistic symptoms.<ref name="Purcel">PMID: 11706102</ref> | [[Autism Spectrum Disorders]] (ASDs) are developmental disorders. During development, glutamate regulates neuronal growth and synaptogenesis, effectively dictating the underlying cytoarchitecture of the brain.<ref name="Purcel"/> Significant research into ASDs has been devoted to understanding how glutamate receptors function and how their disruption might lead to disorders. IGluRs are concentrated in regions of the brain that have been implicated in ASDs including the cerebellum and hippocampus, areas responsible for motor control, spatial navigation and memory. Studies have revealed that glutamate receptor proteins are increased in the cerebellum of autistic individuals by nearly 250% and specific small nucleotide polymorphisms in IGluRs have been identified which correlate with the ASDs. Further, many people with autism have clearly visible disturbances in the anatomy of the inferior olive (IO), a small part of the brain responsible for movement coordination and maintenance of an underlying 12 Hz brain rhythm through careful regulation of glutamate signaling.<ref>PMID: 15749250</ref> A well-known mutation in glutamate receptors in the “Lurcher” mouse model has revealed that constitutively leaky glutamate receptor ion channel result in IO neuron degeneration and loss of purkinje cells, with the mice exhibiting Autism-like characteristics.<ref>PMID: 9285588</ref> Such relationships between overly active glutamate receptors leading to increased excitation/inhibition ratios and autism have led some to propose using glutamate receptor inhibitors as a means of pharmaceutical intervention for improving those with autistic symptoms.<ref>PMID: 14606691</ref> Many pharmacological agents that reduce neural excitation, such as benzodiazapines and anticonvulsants, are thought to potentially have therapeutic value in treating autistic symptoms.<ref name="Purcel">PMID: 11706102</ref> | ||
====GluA2 Structure==== | ====GluA2 Structure==== | ||
| - | AMPA IGluRs form <scene name='Ionotropic_Glutamate_Receptors/Monomer/1'>homotetramers</scene>. Each subunit includes an extracellular <scene name='Ionotropic_Glutamate_Receptors/Atd_opening/1'>amino terminal domain</scene> (ATD) which is responsible for receptor trafficking and modulation, a <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening/1'>ligand-binding domain</scene> (LBD) which activates the receptor upon binding glutamate, and a <scene name='Ionotropic_Glutamate_Receptors/Tmd_opening/1'>transmembrane domain</scene> (TMD) which forms the membrane-spanning ion channel. Also present is a carboxy-terminal domain involved in receptor localization and regulation, although the structure of this domain has not been solved.<ref name="Sobo"/> The structure of AMPA IGluRs or in this case GluA2, is unique in that the <scene name='Ionotropic_Glutamate_Receptors/Sub_a_and_b/1'>symmetry of the receptor changes depending on the domain</scene>. The ATD has a local two-fold symmetry, the LBD has a two-fold symmetry, while the TMD has a four-fold symmetry. Here is a morph depicting the <scene name='Ionotropic_Glutamate_Receptors/Morph_a_to_b/2'>differnce between subunit type A and B</scene>. This symmetry mismatch has implications for function of the receptor with subunits behaving differently depending upon their orientation despite identical primary sequence.<ref name="Sobo"/> For an excellent analysis, see: [[Glutamate_receptor_%28GluA2%29|Glutamate Receptor Symmetry Analysis]] | + | AMPA IGluRs form <scene name='Ionotropic_Glutamate_Receptors/Monomer/1'>homotetramers</scene> composed of <scene name='Ionotropic_Glutamate_Receptors/Distal/1'>distal subunit partners</scene> and <scene name='Ionotropic_Glutamate_Receptors/Proximal/1'>proximal subunits partners</scene>. Each subunit includes an extracellular <scene name='Ionotropic_Glutamate_Receptors/Atd_opening/1'>amino terminal domain</scene> (ATD) which is responsible for receptor trafficking and modulation, a <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening/1'>ligand-binding domain</scene> (LBD) which activates the receptor upon binding glutamate, and a <scene name='Ionotropic_Glutamate_Receptors/Tmd_opening/1'>transmembrane domain</scene> (TMD) which forms the membrane-spanning ion channel. Also present is a carboxy-terminal domain involved in receptor localization and regulation, although the structure of this domain has not been solved.<ref name="Sobo"/> The structure of AMPA IGluRs or in this case GluA2, is unique in that the <scene name='Ionotropic_Glutamate_Receptors/Sub_a_and_b/1'>symmetry of the receptor changes depending on the domain</scene>. The ATD has a local two-fold symmetry, the LBD has a two-fold symmetry, while the TMD has a four-fold symmetry. Here is a morph depicting the <scene name='Ionotropic_Glutamate_Receptors/Morph_a_to_b/2'>differnce between subunit type A and B</scene>. This symmetry mismatch has implications for function of the receptor with subunits behaving differently depending upon their orientation despite identical primary sequence.<ref name="Sobo"/> For an excellent analysis, see: [[Glutamate_receptor_%28GluA2%29|Glutamate Receptor Symmetry Analysis]] |
=====The Amino Terminal Domain===== | =====The Amino Terminal Domain===== | ||
<scene name='Ionotropic_Glutamate_Receptors/Atd_opening/2'>The ATD</scene> is responsible for receptor assembly, trafficking and localization. It has two unique sets of interactions which hold the tetramer together. The <scene name='Ionotropic_Glutamate_Receptors/Atd_dimer_interactions/1'>first set of interactions</scene> is present in each pair of dimers and involves both hydrogen bonding and hydrophobic interactions. The <scene name='Ionotropic_Glutamate_Receptors/Atd_two_dimers_interaction/3'>second set</scene>, which includes residues Ile 203, Thr 204, Ile 205, and Val 209 on both chains among others, effectively holds the pair of dimers together at an angle that is roughly 24 degrees off of the overall two-fold axis.<ref name="Sobo"/><ref>PMID: 19461580</ref> | <scene name='Ionotropic_Glutamate_Receptors/Atd_opening/2'>The ATD</scene> is responsible for receptor assembly, trafficking and localization. It has two unique sets of interactions which hold the tetramer together. The <scene name='Ionotropic_Glutamate_Receptors/Atd_dimer_interactions/1'>first set of interactions</scene> is present in each pair of dimers and involves both hydrogen bonding and hydrophobic interactions. The <scene name='Ionotropic_Glutamate_Receptors/Atd_two_dimers_interaction/3'>second set</scene>, which includes residues Ile 203, Thr 204, Ile 205, and Val 209 on both chains among others, effectively holds the pair of dimers together at an angle that is roughly 24 degrees off of the overall two-fold axis.<ref name="Sobo"/><ref>PMID: 19461580</ref> | ||
| Line 10: | Line 10: | ||
<scene name='Ionotropic_Glutamate_Receptors/Tmd_opening/2'>The TMD</scene> has a pore structure that is nearly identical to that of the [[Potassium Channel]]. With complete four-fold symmetry, 16 helices form a <scene name='Ionotropic_Glutamate_Receptors/Tmd_pore/1'>precise pore</scene> through which cations can flow through. In the current, inhibitor bound structure, the <scene name='Ionotropic_Glutamate_Receptors/Tmd_pore_m3_cross/2'>M3 helices cross</scene> at a highly conserved <scene name='Ionotropic_Glutamate_Receptors/Sytanlaaf_motif/2'>SYTANLAAF motif</scene>, with Thr 617, Ala 621, and Thr 625 occluding the ion permeation pathway.<ref name="Sobo"/> The <scene name='Ionotropic_Glutamate_Receptors/Tmd_narrow/4'>narrowest part</scene> of the channel includes the residues Thr 625, Ala 621, and Thr 617, but does not distinguish between positive cations like in the Potassium Channel. Located next to this narrow region lies <scene name='Ionotropic_Glutamate_Receptors/Tmd_narrow_ala_622/1'>Alanine 622</scene>, which is replaced with a threonine in the Lurcher mouse model mentioned previously. This mutation, which introduces a significantly bulkier residue, destabilizes the tight helix crossing associated with the closed state of the receptor, resulting in a constitutively open ion channel.<ref name="Sobo"/> | <scene name='Ionotropic_Glutamate_Receptors/Tmd_opening/2'>The TMD</scene> has a pore structure that is nearly identical to that of the [[Potassium Channel]]. With complete four-fold symmetry, 16 helices form a <scene name='Ionotropic_Glutamate_Receptors/Tmd_pore/1'>precise pore</scene> through which cations can flow through. In the current, inhibitor bound structure, the <scene name='Ionotropic_Glutamate_Receptors/Tmd_pore_m3_cross/2'>M3 helices cross</scene> at a highly conserved <scene name='Ionotropic_Glutamate_Receptors/Sytanlaaf_motif/2'>SYTANLAAF motif</scene>, with Thr 617, Ala 621, and Thr 625 occluding the ion permeation pathway.<ref name="Sobo"/> The <scene name='Ionotropic_Glutamate_Receptors/Tmd_narrow/4'>narrowest part</scene> of the channel includes the residues Thr 625, Ala 621, and Thr 617, but does not distinguish between positive cations like in the Potassium Channel. Located next to this narrow region lies <scene name='Ionotropic_Glutamate_Receptors/Tmd_narrow_ala_622/1'>Alanine 622</scene>, which is replaced with a threonine in the Lurcher mouse model mentioned previously. This mutation, which introduces a significantly bulkier residue, destabilizes the tight helix crossing associated with the closed state of the receptor, resulting in a constitutively open ion channel.<ref name="Sobo"/> | ||
=====The Ligand Binding Domain===== | =====The Ligand Binding Domain===== | ||
| - | <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening/3'>The LBD</scene> is located just above the TMD and has an overall <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening_two/2'>two-fold axis of symmetry</scene>. Within each LBD lies the so-called <scene name='Ionotropic_Glutamate_Receptors/Lbd_clamshell_open/1'>“clamshell”</scene>. This structure is responsible for <scene name='Ionotropic_Glutamate_Receptors/Lbd_clamshell_open_bound/1'>binding glutamate</scene> and “sensitizing” the receptor to allow passage of cations through the channel. Residues >>>>>>>>>>>>, which are responsible for **tightly binding glutamate** within the clamshell, are highly conserved. Glutamate binding causes a conformational change in the LBD which pulls the M3 helices in the TMD apart, opening the channel and allowing for cation passage. A morph of the conformational change in the LBD upon glutamate binding can be | + | <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening/3'>The LBD</scene> is located just above the TMD and has an overall <scene name='Ionotropic_Glutamate_Receptors/Lbd_opening_two/2'>two-fold axis of symmetry</scene>. Within each LBD lies the so-called <scene name='Ionotropic_Glutamate_Receptors/Lbd_clamshell_open/1'>“clamshell”</scene>. This structure is responsible for <scene name='Ionotropic_Glutamate_Receptors/Lbd_clamshell_open_bound/1'>binding glutamate</scene> and “sensitizing” the receptor to allow passage of cations through the channel. Residues >>>>>>>>>>>>, which are responsible for **tightly binding glutamate** within the clamshell, are highly conserved. Glutamate binding causes a conformational change in the LBD which pulls the M3 helices in the TMD apart, opening the channel and allowing for cation passage. A morph of the conformational change in the LBD upon glutamate binding can be <scene name='Ionotropic_Glutamate_Receptors/Morph_binding/1'>seen here</scene>. Uniquely, due to the varied importance of the homotetramer subunits due to symmetry mismatch, the interaction of glutamate with the distal subunits is predicted to result in a greater conformational change and thus plays a more critical role in channel sensitization and activation.<ref name="Sobo"/> |
====Pharmaceutical Relevance==== | ====Pharmaceutical Relevance==== | ||
As mentioned previously, extensive investigation into the [[Pharmaceutical drugs|pharmaceutical potential]] of IGluRs as a target for treating various ailments including [[Autism Spectrum Disorders]] symptoms is ongoing. In addition to agents which reduce neural excitation such as benzodiazapines and anticonvulsants, small molecules that potentiate AMPA receptor currents have been proven to relieve cognitive deficits caused by neurodegenerative diseases such as [[Alzheimer's Disease]].<ref name="Purcel"/> Modulators such as aniracetam and CX614 **bind on the backside** of the ligand-binding core through interactions with a proline ceiling and a serine floor, stabilizing the closed-clamshell conformation. Although these compounds would likely be ineffective in the case of Autism patients because they slow the deactivation of the IGluR channels, this class of compounds has exciting therapeutic potential.<ref name="Jin"/> | As mentioned previously, extensive investigation into the [[Pharmaceutical drugs|pharmaceutical potential]] of IGluRs as a target for treating various ailments including [[Autism Spectrum Disorders]] symptoms is ongoing. In addition to agents which reduce neural excitation such as benzodiazapines and anticonvulsants, small molecules that potentiate AMPA receptor currents have been proven to relieve cognitive deficits caused by neurodegenerative diseases such as [[Alzheimer's Disease]].<ref name="Purcel"/> Modulators such as aniracetam and CX614 **bind on the backside** of the ligand-binding core through interactions with a proline ceiling and a serine floor, stabilizing the closed-clamshell conformation. Although these compounds would likely be ineffective in the case of Autism patients because they slow the deactivation of the IGluR channels, this class of compounds has exciting therapeutic potential.<ref name="Jin"/> | ||
Revision as of 01:42, 13 March 2011
| |||||||||||
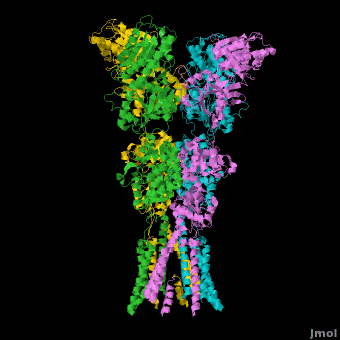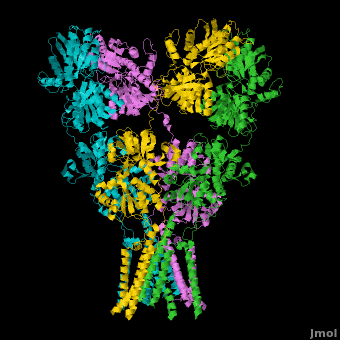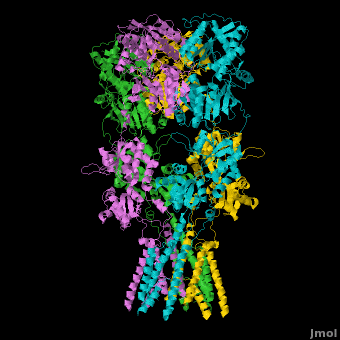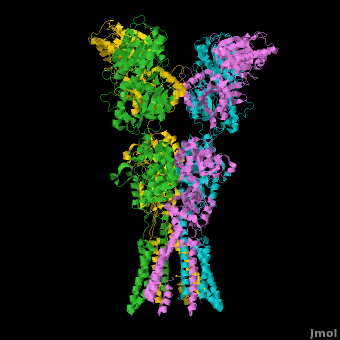

Additional Resources
For additional information on the Symmetry of the Glutamate Receptor, See: Glutamate Receptor Symmetry Analysis
For Additional Information, See: Membrane Channels & Pumps
For Additional Information, See: Alzheimer's Disease
References
- ↑ 1.0 1.1 1.2 Jin R, Clark S, Weeks AM, Dudman JT, Gouaux E, Partin KM. Mechanism of positive allosteric modulators acting on AMPA receptors. J Neurosci. 2005 Sep 28;25(39):9027-36. PMID:16192394 doi:25/39/9027
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 2.6 Sobolevsky AI, Rosconi MP, Gouaux E. X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor. Nature. 2009 Dec 10;462(7274):745-56. Epub . PMID:19946266 doi:10.1038/nature08624
- ↑ 3.0 3.1 3.2 3.3 Purcell AE, Jeon OH, Zimmerman AW, Blue ME, Pevsner J. Postmortem brain abnormalities of the glutamate neurotransmitter system in autism. Neurology. 2001 Nov 13;57(9):1618-28. PMID:11706102
- ↑ Welsh JP, Ahn ES, Placantonakis DG. Is autism due to brain desynchronization? Int J Dev Neurosci. 2005 Apr-May;23(2-3):253-63. PMID:15749250 doi:10.1016/j.ijdevneu.2004.09.002
- ↑ Zuo J, De Jager PL, Takahashi KA, Jiang W, Linden DJ, Heintz N. Neurodegeneration in Lurcher mice caused by mutation in delta2 glutamate receptor gene. Nature. 1997 Aug 21;388(6644):769-73. PMID:9285588 doi:10.1038/42009
- ↑ Rubenstein JL, Merzenich MM. Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003 Oct;2(5):255-67. PMID:14606691
- ↑ Jin R, Singh SK, Gu S, Furukawa H, Sobolevsky AI, Zhou J, Jin Y, Gouaux E. Crystal structure and association behaviour of the GluR2 amino-terminal domain. EMBO J. 2009 Jun 17;28(12):1812-23. Epub 2009 May 21. PMID:19461580 doi:10.1038/emboj.2009.140
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, David Canner, Wayne Decatur, Alexander Berchansky, Joel L. Sussman
