Phosphoglycerate Kinase
From Proteopedia
(Difference between revisions)
| Line 16: | Line 16: | ||
== Reaction Mechanism == | == Reaction Mechanism == | ||
| - | The bilobed structure of PGK is very crucial in its catalytic function. The active site is broken into two pieces, one on the interior of each lobe or domain. The N-terminal domain has a basic region where the 1,3-Biphosphoglycerate and 3-phosphoglycerate bind while the C-terminal domain has the binding sites for the nucleotide substrates, ADP and ATP. Upon binding of both substrate molecules at the active sites, the protein’s conformation changes such that the two lobes of the protein swing together <ref>Voet, Donald et al. 2008. Fundamentals of Biochemistry. 3rd ed. 499 </ref> When the two domains swing shut, a hydrophobic chamber free from water is established where the reaction can take place. This hydrophobic chamber is necessary to prevent ATP hydrolysis <ref name="Auer" /> The hinge for this conformational change is beta sheet L and the new conformation is formed via a salt bridge between <scene name='Shane_Harmon_Sandbox/Arg_and_asp/2'>Arg 62 and Asp 200</scene> <ref>PMID:6115427</ref> Recent research indicates that the mechanism for closure of the two domains is a series of hydrogen bond interactions that occur upon binding of the substrates on both domains | + | The bilobed structure of PGK is very crucial in its catalytic function. The active site is broken into two pieces, one on the interior of each lobe or domain. The N-terminal domain has a basic region where the 1,3-Biphosphoglycerate and 3-phosphoglycerate bind while the C-terminal domain has the binding sites for the nucleotide substrates, ADP and ATP. Upon binding of both substrate molecules at the active sites, the protein’s conformation changes such that the two lobes of the protein swing together <ref>Voet, Donald et al. 2008. Fundamentals of Biochemistry. 3rd ed. 499 </ref> When the two domains swing shut, a hydrophobic chamber free from water is established where the reaction can take place. This hydrophobic chamber is necessary to prevent ATP hydrolysis <ref name="Auer" /> The hinge for this conformational change is beta sheet L and the new conformation is formed via a salt bridge between <scene name='Shane_Harmon_Sandbox/Arg_and_asp/2'>Arg 62 and Asp 200</scene> <ref>PMID:6115427</ref> Recent research indicates that the mechanism for closure of the two domains is a series of hydrogen bond interactions that occur upon binding of the substrates on both domains <ref>PMID:20088776 </ref> |
The mechanism of catalysis has not been fully established because the PGK/1-3biphophoglycerate complex is highly unstable; however, it is thought that the mechanism is similar to that of hexokinase. Hexokinase catalyzes the removal of a phosphate group from ATP to glucose and has a very similar structure and conformational change via a hinge. PGK has a similar function except it catalyzes the transfer of a phosphate to form ATP instead of using ATP. The reaction of PGK removes the C1 phosphate group from 1,3-biphosphoglycerate and transfers it to ADP to form ATP. Once the substrates bind to the active sites, the protein domains swing shut forcing the substrates into correct position for the reaction to proceed <ref>Harnan, G. et al. 1992. Domain Motions in Phosphoglycerate Kinase: Determination of Interdomain Distance Distribution by Site Specific Labeling and Time Resolved Flourescense Energy Transfer. PNAS. 89:11764-11768.</ref> | The mechanism of catalysis has not been fully established because the PGK/1-3biphophoglycerate complex is highly unstable; however, it is thought that the mechanism is similar to that of hexokinase. Hexokinase catalyzes the removal of a phosphate group from ATP to glucose and has a very similar structure and conformational change via a hinge. PGK has a similar function except it catalyzes the transfer of a phosphate to form ATP instead of using ATP. The reaction of PGK removes the C1 phosphate group from 1,3-biphosphoglycerate and transfers it to ADP to form ATP. Once the substrates bind to the active sites, the protein domains swing shut forcing the substrates into correct position for the reaction to proceed <ref>Harnan, G. et al. 1992. Domain Motions in Phosphoglycerate Kinase: Determination of Interdomain Distance Distribution by Site Specific Labeling and Time Resolved Flourescense Energy Transfer. PNAS. 89:11764-11768.</ref> | ||
Revision as of 09:51, 24 August 2018
| |||||||||||
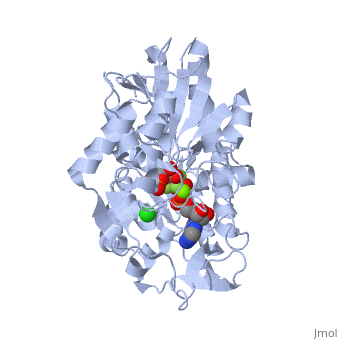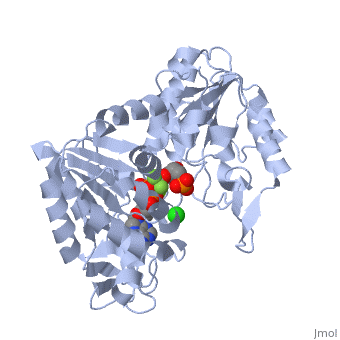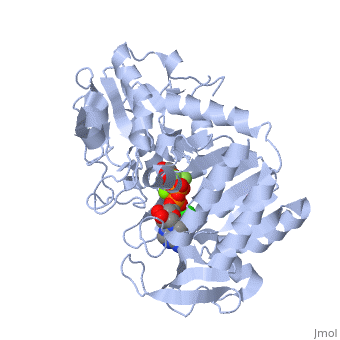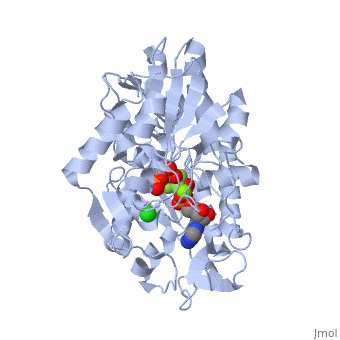

3D structures of phosphoglycerate kinase
Updated on 24-August-2018
Additional Resources
For additional information, see: Carbohydrate Metabolism
References
- ↑ 1.0 1.1 1.2 1.3 Auerbach G, Huber R, Grattinger M, Zaiss K, Schurig H, Jaenicke R, Jacob U. Closed structure of phosphoglycerate kinase from Thermotoga maritima reveals the catalytic mechanism and determinants of thermal stability. Structure. 1997 Nov 15;5(11):1475-83. PMID:9384563
- ↑ Lallemand P, Chaloin L, Roy B, Barman T, Bowler MW, Lionne C. Interaction of human 3-phosphoglycerate kinase with its two substrates: is substrate antagonism a kinetic advantage? J Mol Biol. 2011 Jun 24;409(5):742-57. Epub 2011 Apr 27. PMID:21549713 doi:10.1016/j.jmb.2011.04.048
- ↑ Voet, Donald et al. 2008. Fundamentals of Biochemistry. 3rd ed. 499
- ↑ Blake CC, Rice DW. Phosphoglycerate kinase. Philos Trans R Soc Lond B Biol Sci. 1981 Jun 26;293(1063):93-104. PMID:6115427
- ↑ Vas M, Varga A, Graczer E. Insight into the Mechanism of Domain Movements and their Role in Enzyme Function: Example of 3-Phosphoglycerate Kinase. Curr Protein Pept Sci. 2010 Jan 21. PMID:20088776
- ↑ Harnan, G. et al. 1992. Domain Motions in Phosphoglycerate Kinase: Determination of Interdomain Distance Distribution by Site Specific Labeling and Time Resolved Flourescense Energy Transfer. PNAS. 89:11764-11768.
- ↑ Scopes, Robert. 1977. The Steady State Kinetics of Yeast Phosphoglycerate Kinase. European Journal of Biochemistry. 85, 503-516
- ↑ Macioszek, Jerzy et al. 1990. Kinetics of the Two-Enzyme Phosphoglycerate Kinase/Glyceraldehyde-3-Phosphate Dehydrogenase Couple. Plant Physiology 94: 291-296.
- ↑ Shaobo, Wu et al. 2009. PGK1 expression responds to freezing, anoxia, and dehydration stresses in freeze tolerant wood frog, Rana sylvatica. Journal of Experimental Zoology. 311, 57-67
- ↑ Hogg, PJ. 2002. Biological Regulation through protein disulfide bond cleavage. Redox Report. 7(2), 71-77.
Proteopedia Page Contributors and Editors (what is this?)
Shane Harmon, Michal Harel, Joel L. Sussman, Brandon Tritle, David Canner, Alexander Berchansky
