Gossypium hirsutum gene of unknown function Gohir.A03G0737001 encodes a potential Chaperone-like Protein of protochlorophyllide oxidoreductase (CPP1)
Alana N. Osborne, Andrew Osagiede, Amanda R. Storm, Amanda M. Hulse-Kemp, Angela K. Stoeckman [1]
Molecular Tour
Cotton plays a significant role in the United States economy and research on cotton is essential as the price of cotton and the financial gain to the cotton grower is dependent on the fiber yield and quality. Recently, the genomes of five allotetraploid cotton species (Gossypium hirsutum, Gossypium barbadense, Gossypium mustelinum, Gossypium tomentosum, Gossypium darwinii) were sequenced[2], and analysis of gene annotations revealed many genes of unknown function in common across the species. One of these genes of unknown function, the Gossypium hirsutum gene Gohir.A03G073700 and associated protein (UniProt: A0A1U8HKT6) here referred to as GhCPP1-A0A1U8HKT6, was studied using bioinformatic tools and is proposed to be a chaperone-like protein of POR1 (CPP1).
Most plants require light for the biosynthesis of the green pigment chlorophyll, which is central to the process of photosynthesis. One of the later steps of the chlorophyll biosynthetic pathway is facilitated by protochlorophyllide oxidoreductase (POR), a light-dependent enzyme[3]. To complete chlorophyll biosynthesis, the POR enzyme must be post-translationally imported into the chloroplast. The chaperone-like protein of POR1 (CPP1) in Arabidopsis has been shown to regulate POR transport and function during this process although its specific role in the process is still unclear[3]. Due to the structural similarity between the J-like domain in CPP1 and the J domain in the DnaJ/HSP70 protein family, it has been suggested that CPP1 has a function similar to the “holdase” chaperone function of the J domain that helps organize interactions with Hsp70 chaperone partners[4]. A functionally conserved His-Pro-Asp (HPD) motif within J-domains facilitates this interaction and, although very similar in structure to the J domain, the HPD motif is absent in J-like domain proteins such as CPP1, indicating these domains may perform a similar molecular function but may have diverged interaction partners.
Sequence analysis of GhCPP1-A0A1U8HKT6 indicated it has a N-terminal chloroplast transit peptide with a cleavage site at amino acid 45 along with four predicted TM regions in the C-terminal half of the protein. A domain architecture was created to visualize these sequence features (see static image below):
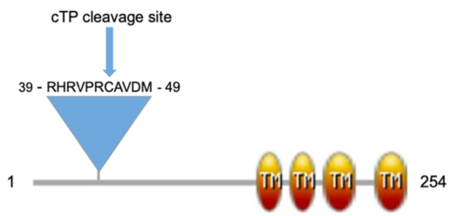
Domain architecture of GhCPP1-A0A1U8HKT6 created using Prosite:MyDomains (Hulo et al. 2008) based on predictions from InterPro (Blum et al. 2021), TM indicates transmembrane, cTP is chloroplast transit peptide.
Interestingly, Arabidopsis thaliana and Nicotiana benthamiana CPP1 proteins are indicated to contain three, not four, TM regions [3]. However, a homolog of Arabidopsis CPP1, At2g20920, has been identified to also contain four TM regions[5] suggesting a potential structural or functional difference between these family members.
The (colored in rainbow format with the N-terminus in blue and C-terminus in red) depicts between amino acids 56-119 and 150-254 and a low-confidence modeled region between them from amino acids 119-150 (shown transparent). The predicted TM regions are seen as a 4-helix bundle that exhibits expected (Hydrophobic, Polar). (PDB ID: 2dn9; in deep sky blue) displays a similar anti-parallel hairpin between the horizontally-displayed alpha helices II and III, although the GhCPP1-A0A1U8HKT6 J-like domain lacks the fourth helix of human Tid1 and has a shorter loop.
When DALI [6] was used to search for proteins having a similar tertiary structure to the proposed J-like domain of GhCPP1-A0A1U8HKT6, one of the highest similarity hits (Z-score of 7.9 and RMSD score of 1.5) was with the J-like domain of yeast TIM16 that is part of the mitochondrial translocase of the inner membrane protein complex, TIM16/TIM14 (PDB ID: 2guz). The TIM16/TIM14 heterodimer complex serves as a co-chaperone duo, a critical part of the translocase import motor responsible for piloting translocation proteins into the mitochondria[7]. As the biosynthesis of chlorophyll in plants requires the light-dependent enzyme POR1 to be post-translationally imported into the chloroplast, it is possible that the J-like domain found in GhCPP1-A0A1U8HKT6 may allow it to partner with an as yet unknown co-chaperone to function similarly to the TIM16/TIM14 complex, chaperoning the translocation of POR1. This is supported by an overlay of the GhCPP1-A0A1U8HKT6 J-like domain (transparent ConSurf colored surface) with the J-like domain of TIM16 (light blue ribbon, PDB 2guz) that shows a conserved surface on GhCPP1-A0A1U8HKT6 at the modeled protein-protein interaction site with the TIM14 protein (dark blue ribbon).
Evidence from protein sequence, homology, and structure support GhCPP1-A0A1U8HKT6 as a chaperone-like protein of POR1 in Gossypium hirsutum. GhCPP1-A0A1U8HKT6 is predicted to be anchored in the chloroplast membrane where it could interact with POR and support its entry into the chloroplast where chlorophyll biosynthesis occurs, similar to Arabidopsis CPP1 to which it has high sequence similarity. The molecular function of the J-like domain is still unclear. Similarity in structure to the J-domain suggests a similar protein-protein interaction domain function as in the “holdase” activity of molecular chaperones, although the lack of a HPD motif indicates different binding partners. This is supported by the J-like domain in mitochondrial TIM16 being part of a heterodimer complex. It would be interesting for further studies to explore potential binding partners of GhCPP1-A0A1U8HKT6.
References
- ↑ doi: https://dx.doi.org/10.17912/micropub.biology.000867
- ↑ Chen ZJ, Sreedasyam A, Ando A, Song Q, De Santiago LM, Hulse-Kemp AM, Ding M, Ye W, Kirkbride RC, Jenkins J, Plott C, Lovell J, Lin YM, Vaughn R, Liu B, Simpson S, Scheffler BE, Wen L, Saski CA, Grover CE, Hu G, Conover JL, Carlson JW, Shu S, Boston LB, Williams M, Peterson DG, McGee K, Jones DC, Wendel JF, Stelly DM, Grimwood J, Schmutz J. Genomic diversifications of five Gossypium allopolyploid species and their impact on cotton improvement. Nat Genet. 2020 May;52(5):525-533. doi: 10.1038/s41588-020-0614-5. Epub 2020 Apr , 20. PMID:32313247 doi:http://dx.doi.org/10.1038/s41588-020-0614-5
- ↑ 3.0 3.1 3.2 Lee JY, Lee HS, Song JY, Jung YJ, Reinbothe S, Park YI, Lee SY, Pai HS. Cell growth defect factor1/chaperone-like protein of POR1 plays a role in stabilization of light-dependent protochlorophyllide oxidoreductase in Nicotiana benthamiana and Arabidopsis. Plant Cell. 2013 Oct;25(10):3944-60. PMID:24151298 doi:10.1105/tpc.113.111096
- ↑ Kelley WL. The J-domain family and the recruitment of chaperone power. Trends Biochem Sci. 1998 Jun;23(6):222-7. PMID:9644977 doi:10.1016/s0968-0004(98)01215-8
- ↑ Kawai-Yamada M, Nagano M, Kakimoto M, Uchimiya H. Plastidic protein Cdf1 is essential in Arabidopsis embryogenesis. Planta. 2014 Jan;239(1):39-46. PMID:24097264 doi:10.1007/s00425-013-1966-1
- ↑ Holm L. Using Dali for Protein Structure Comparison. Methods Mol Biol. 2020;2112:29-42. doi: 10.1007/978-1-0716-0270-6_3. PMID:32006276 doi:http://dx.doi.org/10.1007/978-1-0716-0270-6_3
- ↑ Mokranjac D, Bourenkov G, Hell K, Neupert W, Groll M. Structure and function of Tim14 and Tim16, the J and J-like components of the mitochondrial protein import motor. EMBO J. 2006 Oct 4;25(19):4675-85. Epub 2006 Sep 14. PMID:16977310


