Figure 1.
 }
}
Mechanism
Polyethylene terephthalate (PET) hydrolase (PETase) is a serine hydrolase that catalyzes the cleavage of ester bonds in PET polymers. 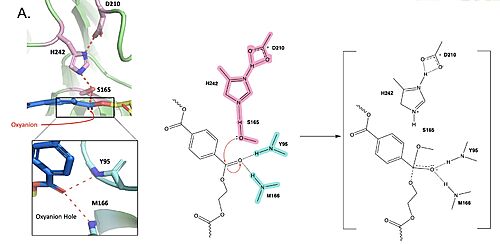
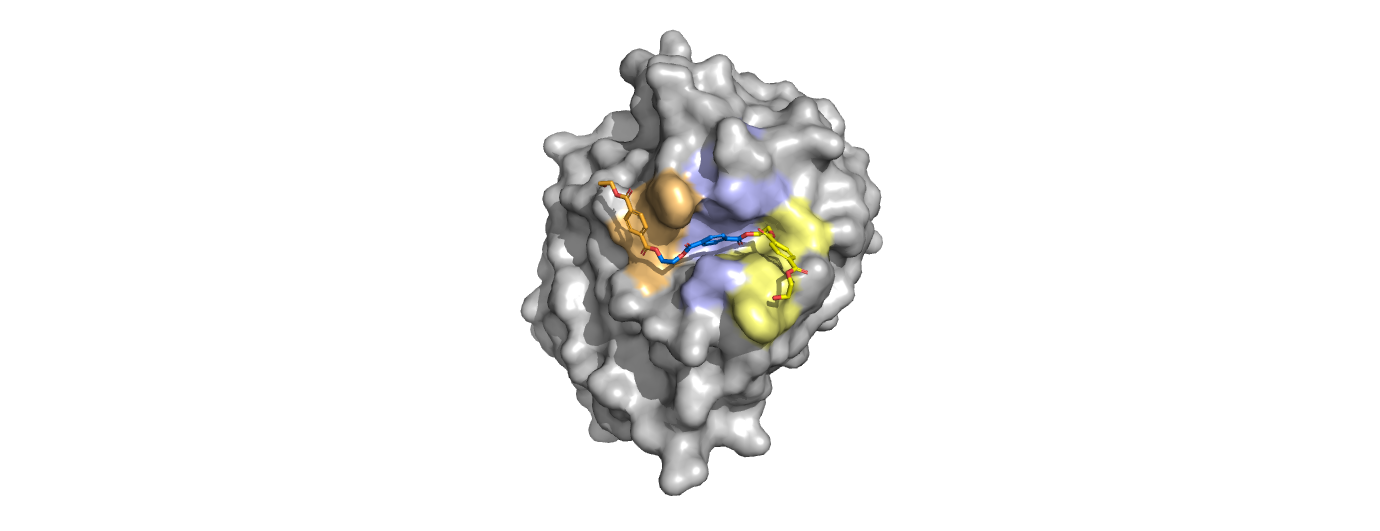
Figure 1. Active site of PETase highlighting catalytic residues and the first step of the reaction mechanism. Residues in pink represent the catalytic triad (S165, H242, and D210). Light blue represents the oxyanion hole residues (Y95 and M166).
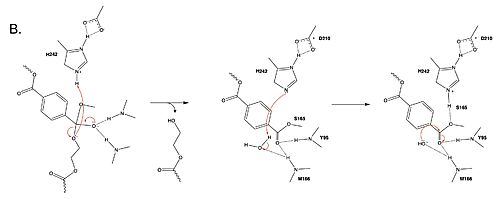
Figure 2. Second step of the PET hydrolysis reaction by PETase.
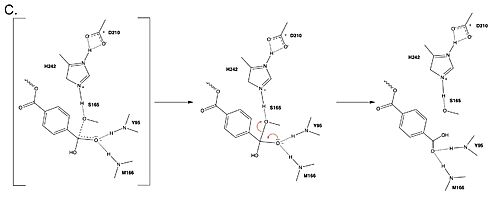
Figure 3. Final step of PET hydrolysis by PETase.
Mutations
Researchers have been investigating various mutations of PET hydrolase to enhance its catalytic ability. One group of researchers, Tournier et. al., have made mutations in the PET hydrolase active site. They identified the key residues involved in the catalytic mechanism by using a model of the (2-HE(MHET)₃) onto the enzyme (PDB ID 4EB0). The site, mainly a hydrophobic pocket, contained 11 residues targeted for mutagenesis. From this, they identified that the majority of enzymes' specific activity went down; however, the mutation of the to either isoleucine or tryptophan increased specific activity. Four target mutations introduced into the PET Hydrolase by Tournier et. al. demonstrated improved catalytic efficiency and thermal stability compared to its wild-type structure.
F243W Mutation
The mutation of the F243 position to a tryptophan () was selected for further analysis based on its enhanced catalytic activity in the depolymerization of Pf-PET. The W243 mutation was shown to improve the substrate's binding affinity and increase the enzyme’s activity compared to the wild-type enzyme. This was one of the few variants that exhibited higher activity, and it was further analyzed through differential scanning fluorimetry (DSF) to assess its stability. [1]
F243I Mutation
Similar to the W243 mutation, the was identified as a variant with improved depolymerization activity of Pf-PET. The I243 mutation in LCC led to better substrate interaction than the wild-type enzyme. After generating all possible variants, this mutation was among the few that exhibited 75% or more of the wild-type specific activity. As with the W243 mutation, DSF was used to determine the melting temperature and thermal stability, supporting the increased activity observed with this mutation. [1]
F243W/I Results
Table 1. Specific activity and rate of wild-type, WCCG, and ICCG mutants. Initial rate was measured by the calculated rate of reaction by NaOH consumption. Specific activity was measured via pf-PET-depolymerization assay. [1]
| Enzyme |
| Initial rate (ghydrolyzed PET•L-1•h-1) |
| Specific activity (mgTAeqh-1•mgenzyme-1) ± SD |
| Wild-Type |
| 25.7 |
| 81.9 ± 5.6 |
| WCCG |
| 30.3 |
| 75.9 ± 5.9 |
| ICCG |
| 31.0 |
| 82.0 ± 3.9 |
Both the WCCG and ICCG mutants display slightly higher initial rates (30.3 and 31.0 g hydrolyzed PET•L⁻¹•h⁻¹, respectively) compared to the wild-type enzyme (25.7 g hydrolyzed PET•L⁻¹•h⁻¹). This suggests that the mutations introduced in WCCG and ICCG enhance the rate of PET breakdown.
The specific activity of the WCCG mutant (75.9 mg TAeq h⁻¹ mg⁻¹ enzyme) is slightly lower than that of wild-type PETase (81.9 mg TAeq h⁻¹ mg⁻¹ enzyme), whereas the ICCG mutant shows comparable specific activity to the wild-type (82.0 mg TAeq h⁻¹ mg⁻¹ enzyme). Mutating the active site F243 to I243 (in the ICCG mutant) and W243 (in the WCCG mutant) resulted in a percent gain of specific activity of 127.5% and 118.4% respectively. [1]
D238C/S283C Mutations
A disulfide bond was put into the structure of the LCC by mutating into a . The disulfide bond specifically replaced those residues due to the triplet acidic residue sequence E208/D238/S283 serving as a common divalent metal binding site in other cutinase enzymes. [1] This allowed the researchers to compare the activity of the disulfide bond mutation’s activity and stability to that of the cutinases with additional metal ions that showed increased activity and stability. (Disulfide bonds are also comparable to metal cations in terms of enhancing protein structural thermal stability.) The disulfide bond proved true to enhance the thermal stability of the PET hydrolase, increasing its melting temperature to 94.5 degrees Celsius to that of the wild-type’s 84.7 degree Celsius thermal stability. [1] A tradeoff with catalytic activity occurred, though, decreasing enzymatic activity by 28% compared to the wild type. The thermal stability seemed to be more important than activity rates if the LCC still did its intended function. [1]
Y127G Mutation
A was introduced to residue the . The main effect of this mutation was that it showed an increase in melting temperature, but with no effect on depolymerization of the PET substrates compared to the wild type. [1] With the addition of the glycine, the ICC/WCC mutations showed a similar or higher specific activity with a higher melting point compared to the wild type (9.3 to 13.4 degrees Celsius higher). [1] Distances between the catalytic serine (S165) and the ligand where the scissile bond is located, as well as with the catalytic histidine (H242), decreased to enhance catalytic efficiency of LCC with new covalent interactions facilitated. [1]
Glycosylation
were introduced in a research study completed by Abhihit N. Shirke and others with the initial intention of inducing aggregation in the leaf branch cutinase / PET hydrolase wild-type. [3] Glycosylation, as a general tool, is introduced into a protein to improve conformational stability. The specific type used in this study was N-linked side chain alteration. This means that the glycosylation sites were selected based on a starting asparagine residue followed by the sequence N-X-S or N-X-T, where X stands for any of the twenty amino acids except proline. [6] This is implemented specifically because it allows for a better ability to choose the mutation sites. The first glycosylation site followed the N-T-S pattern with . The second was with an N-A-S pattern, located nearest to the active site. The final was , exhibiting a N-D-T sequence. [3] With the combination of these glycosylation sites (without any other mutagenesis introduced to the enzyme), an overall 10-degree Celsius higher thermal stability was exhibited compared to the wild-type, with the structure of the modified PET hydrolase three times more stable. Catalytic efficiency also improved at the enzyme's known melting temperature. Even though the target point of introducing glycosylation sites was to induce aggregation in the PET hydrolase, depletion of aggregation was exhibited. The glycosylated protein took twice as long to unfold compared to the wild-type. For comparison, the threshold of 80 degrees Celsius was where the major difference in kinetic activity occurred: 85% of the glycosylated hydrolase maintained its catalytic activity, whereas the wild-type only had 50% of it working at the same temperature. [3] The first and third sites showed these trends both together and on their own as the only glycosylation sites, but the second one nearest the active site showed no change when glycosylated on its own when compared to the wild type. [3]
Biochemical Results
Improved Thermal Stability
Thermal stability is very important for enzyme-catalyzed PET degradation because the reaction must take place above the transition temperature of PET (70ºC), which allows the substrate to have optimal flexibility to fit into the active site. The disulfide bridge mutation raises the melting point of the enzyme from 84.7ºC to 94.5ºC. [1] The covalent disulfide bond allows the tertiary structure of the enzyme to maintain its integrity at higher temperatures.
Depolymerization Efficiency of Mutant LCCs
Both the ICCG and WCCG mutants designed by Tournier et al. exhibited a greater depolymerization efficiency than the wild type. The wild type LCC reached 53% depolymerization in 20 hours while both the ICCG and WCCG mutants reached 90% depolymerization in 10.5 hrs and 9.3 hrs respectively. [1]
Enhanced Catalytic Efficiency
The ICCG and WCCG mutations constructed by Tournier et al. showed a return to the wild type activity and beyond. The WCCG quadruple mutation showed a 122% increase in catalysis, with an increased ability to sustain its structure at temperatures 6.2 degrees higher than the wild type (84.7 degrees Celsius). [1] The ICCG quadruple mutation showed a 2% decrease in activity compared to the wild type, but a thermal stability increase by 10.1 degrees Celsius. [1] The increase in specific activity of the F243W mutation could be attributed to the hydrogen bond formed between the amine group on tryptophan and a carbonyl oxygen of the -1 monomer, with the aromatic ring in W243 retaining pi-stacking and Van der Waals interactions made by F243 in the wild type. These additional intermolecular forces between the active site and the ligand likely led to improved binding affinity which increased specific activity.
Conclusions
Implications for Plastic Recycling
Enzyme-catalyzed plastic degradation produces less sodium sulfate NaSO4 than other recycling processes. Current standard recycling processes produce approximately 80% NaSO4 by weight of recycled material. [7] There are many negative environmental impacts of NaSO4 including acid rain, respiratory health concerns, reduced visibility due to light refraction by atmospheric particles, and disrupting aquatic ecosystems. Therefore, it is essential to mitigate NaSO4 waste produced in recycling processes.
This improved enzyme-catalyzed mechanism of PET depolymerization has important implications in horizontal recycling–the process of recycling used products back into their raw materials to create the same or similar products [8], and could help to close the loop of the circular economy. The LCC mutant engineered by Tournier et al. is a promising aid in the issue of excessive plastic disposal.




