This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
1j3h
From Proteopedia
(New page: 200px<br /><applet load="1j3h" size="450" color="white" frame="true" align="right" spinBox="true" caption="1j3h, resolution 2.9Å" /> '''Crystal structure of ...) |
|||
| Line 1: | Line 1: | ||
| - | [[Image:1j3h.gif|left|200px]]<br /><applet load="1j3h" size=" | + | [[Image:1j3h.gif|left|200px]]<br /><applet load="1j3h" size="350" color="white" frame="true" align="right" spinBox="true" |
caption="1j3h, resolution 2.9Å" /> | caption="1j3h, resolution 2.9Å" /> | ||
'''Crystal structure of apoenzyme cAMP-dependent protein kinase catalytic subunit'''<br /> | '''Crystal structure of apoenzyme cAMP-dependent protein kinase catalytic subunit'''<br /> | ||
==Overview== | ==Overview== | ||
| - | To better understand the mechanism of ligand binding and ligand-induced | + | To better understand the mechanism of ligand binding and ligand-induced conformational change, the crystal structure of apoenzyme catalytic (C) subunit of adenosine-3',5'-cyclic monophosphate (cAMP)-dependent protein kinase (PKA) was solved. The apoenzyme structure (Apo) provides a snapshot of the enzyme in the first step of the catalytic cycle, and in this unliganded form the PKA C subunit adopts an open conformation. A hydrophobic junction is formed by residues from the small and large lobes that come into close contact. This "greasy" patch may lubricate the shearing motion associated with domain rotation, and the opening and closing of the active-site cleft. Although Apo appears to be quite dynamic, many important residues for MgATP binding and phosphoryl transfer in the active site are preformed. Residues around the adenine ring of ATP and residues involved in phosphoryl transfer from the large lobe are mostly preformed, whereas residues involved in ribose binding and in the Gly-rich loop are not. Prior to ligand binding, Lys72 and the C-terminal tail, two important ATP-binding elements are also disordered. The surface created in the active site is contoured to bind ATP, but not GTP, and appears to be held in place by a stable hydrophobic core, which includes helices C, E, and F, and beta strand 6. This core seems to provide a network for communicating from the active site, where nucleotide binds, to the peripheral peptide-binding F-to-G helix loop, exemplified by Phe239. Two potential lines of communication are the D helix and the F helix. The conserved Trp222-Phe238 network, which lies adjacent to the F-to-G helix loop, suggests that this network would exist in other protein kinases and may be a conserved means of communicating ATP binding from the active site to the distal peptide-binding ledge. |
==About this Structure== | ==About this Structure== | ||
| - | 1J3H is a [http://en.wikipedia.org/wiki/Single_protein Single protein] structure of sequence from [http://en.wikipedia.org/wiki/Mus_musculus Mus musculus] with MPD as [http://en.wikipedia.org/wiki/ligand ligand]. Active as [http://en.wikipedia.org/wiki/Non-specific_serine/threonine_protein_kinase Non-specific serine/threonine protein kinase], with EC number [http://www.brenda-enzymes.info/php/result_flat.php4?ecno=2.7.11.1 2.7.11.1] Full crystallographic information is available from [http:// | + | 1J3H is a [http://en.wikipedia.org/wiki/Single_protein Single protein] structure of sequence from [http://en.wikipedia.org/wiki/Mus_musculus Mus musculus] with <scene name='pdbligand=MPD:'>MPD</scene> as [http://en.wikipedia.org/wiki/ligand ligand]. Active as [http://en.wikipedia.org/wiki/Non-specific_serine/threonine_protein_kinase Non-specific serine/threonine protein kinase], with EC number [http://www.brenda-enzymes.info/php/result_flat.php4?ecno=2.7.11.1 2.7.11.1] Full crystallographic information is available from [http://oca.weizmann.ac.il/oca-bin/ocashort?id=1J3H OCA]. |
==Reference== | ==Reference== | ||
| Line 15: | Line 15: | ||
[[Category: Single protein]] | [[Category: Single protein]] | ||
[[Category: Akamine, P.]] | [[Category: Akamine, P.]] | ||
| - | [[Category: Eyck, L | + | [[Category: Eyck, L F.Ten.]] |
[[Category: Madhusudan]] | [[Category: Madhusudan]] | ||
| - | [[Category: Taylor, S | + | [[Category: Taylor, S S.]] |
[[Category: Wu, J.]] | [[Category: Wu, J.]] | ||
| - | [[Category: Xuong, N | + | [[Category: Xuong, N H.]] |
[[Category: MPD]] | [[Category: MPD]] | ||
[[Category: apoenzyme]] | [[Category: apoenzyme]] | ||
| Line 28: | Line 28: | ||
[[Category: preformed active site]] | [[Category: preformed active site]] | ||
| - | ''Page seeded by [http:// | + | ''Page seeded by [http://oca.weizmann.ac.il/oca OCA ] on Thu Feb 21 13:18:31 2008'' |
Revision as of 11:18, 21 February 2008
|
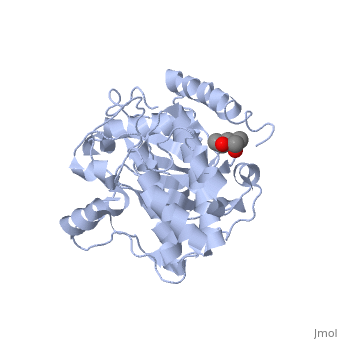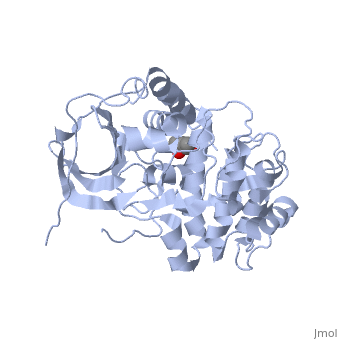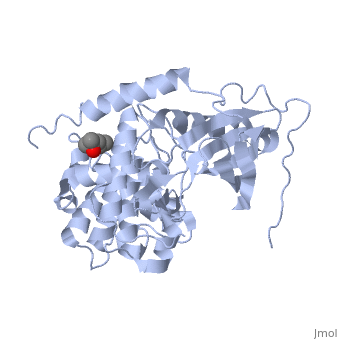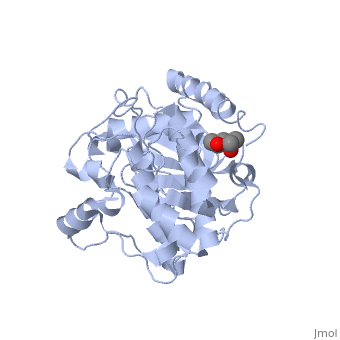
Crystal structure of apoenzyme cAMP-dependent protein kinase catalytic subunit
Overview
To better understand the mechanism of ligand binding and ligand-induced conformational change, the crystal structure of apoenzyme catalytic (C) subunit of adenosine-3',5'-cyclic monophosphate (cAMP)-dependent protein kinase (PKA) was solved. The apoenzyme structure (Apo) provides a snapshot of the enzyme in the first step of the catalytic cycle, and in this unliganded form the PKA C subunit adopts an open conformation. A hydrophobic junction is formed by residues from the small and large lobes that come into close contact. This "greasy" patch may lubricate the shearing motion associated with domain rotation, and the opening and closing of the active-site cleft. Although Apo appears to be quite dynamic, many important residues for MgATP binding and phosphoryl transfer in the active site are preformed. Residues around the adenine ring of ATP and residues involved in phosphoryl transfer from the large lobe are mostly preformed, whereas residues involved in ribose binding and in the Gly-rich loop are not. Prior to ligand binding, Lys72 and the C-terminal tail, two important ATP-binding elements are also disordered. The surface created in the active site is contoured to bind ATP, but not GTP, and appears to be held in place by a stable hydrophobic core, which includes helices C, E, and F, and beta strand 6. This core seems to provide a network for communicating from the active site, where nucleotide binds, to the peripheral peptide-binding F-to-G helix loop, exemplified by Phe239. Two potential lines of communication are the D helix and the F helix. The conserved Trp222-Phe238 network, which lies adjacent to the F-to-G helix loop, suggests that this network would exist in other protein kinases and may be a conserved means of communicating ATP binding from the active site to the distal peptide-binding ledge.
About this Structure
1J3H is a Single protein structure of sequence from Mus musculus with as ligand. Active as Non-specific serine/threonine protein kinase, with EC number 2.7.11.1 Full crystallographic information is available from OCA.
Reference
Dynamic features of cAMP-dependent protein kinase revealed by apoenzyme crystal structure., Akamine P, Madhusudan, Wu J, Xuong NH, Ten Eyck LF, Taylor SS, J Mol Biol. 2003 Mar 14;327(1):159-71. PMID:12614615
Page seeded by OCA on Thu Feb 21 13:18:31 2008
Categories: Mus musculus | Non-specific serine/threonine protein kinase | Single protein | Akamine, P. | Eyck, L F.Ten. | Madhusudan | Taylor, S S. | Wu, J. | Xuong, N H. | MPD | Apoenzyme | Camp-dependent protein kinase | Catalytic subunit | Crystal structure | Open conformation | Preformed active site

{kind=link}
{kind=link}