This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
User:Eran Hodis/Sandbox3
From Proteopedia
(→Your Heading Here (maybe something like 'Structure')) |
|||
| Line 4: | Line 4: | ||
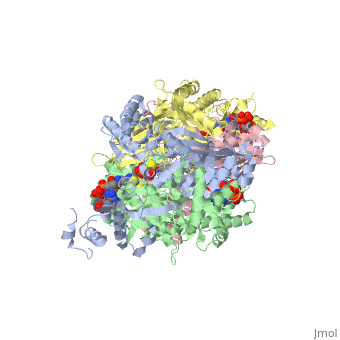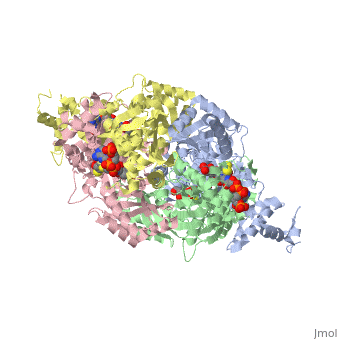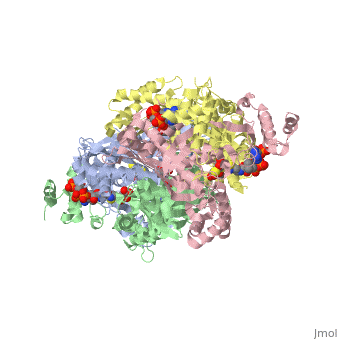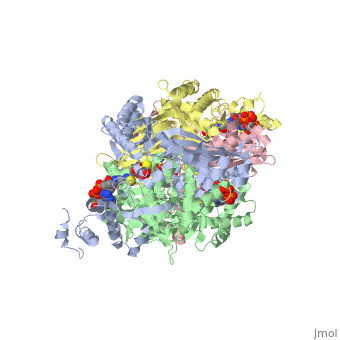
There are two distinct classes of HMGRs, class I, which is only found in eukaryotes and are membrane bound and class II, which is found in prokaryotes and are soluble. <ref>PMID:11349148</ref> HMGR contains 8 transmembrane domains that have yet to be successfully crystallized, which anchor the protein to the membrane of the endoplasmic reticulum. <ref name="Roitelman"/> The catalytic portion of human HMGR forms a tetramer, with the individual monomers winding around each other. <ref name="Roitelman">PMID:1374417</ref> Within the tetramer, the monomers are arranged into <scene name='HMG-CoA_Reductase/1dq8_2_dimers/3'>two dimers</scene>, each of which contains <scene name='HMG-CoA_Reductase/1dq8_2_active_sites/2'>two active sites </scene>which are formed by residues form both monomers. Each monomer contains <scene name='HMG-CoA_Reductase/1dq8_star3_domains/2'>three domains </scene>, the <scene name='HMG-CoA_Reductase/1dq8_n_domain/2'>N-domain</scene>, the <scene name='HMG-CoA_Reductase/1dq8_l_domain/1'>L-Domain</scene>, and the <scene name='HMG-CoA_Reductase/1dq8_s_domain/1'>S-Domain</scene>. The L-domain is unique to HMGRs while the S-domain, which forms the binding site for NADP, resembles that of [[ferredoxin]]. The S and L domains are connected by a <scene name='HMG-CoA_Reductase/1dq8_cis_loop/6'>“cis-loop”</scene> which is essential for the HMG-binding site. <ref name="Roitelman"/> Salt bridges between residues R641 and E782 as well as <scene name='HMG-CoA_Reductase/1dq8_cis_loop/4'>hydrogen bonds</scene> between E700 and E700 on neighboring monomers compliment the largely hydrophobic dimer-dimer interface. <ref name="Roitelman"/> | There are two distinct classes of HMGRs, class I, which is only found in eukaryotes and are membrane bound and class II, which is found in prokaryotes and are soluble. <ref>PMID:11349148</ref> HMGR contains 8 transmembrane domains that have yet to be successfully crystallized, which anchor the protein to the membrane of the endoplasmic reticulum. <ref name="Roitelman"/> The catalytic portion of human HMGR forms a tetramer, with the individual monomers winding around each other. <ref name="Roitelman">PMID:1374417</ref> Within the tetramer, the monomers are arranged into <scene name='HMG-CoA_Reductase/1dq8_2_dimers/3'>two dimers</scene>, each of which contains <scene name='HMG-CoA_Reductase/1dq8_2_active_sites/2'>two active sites </scene>which are formed by residues form both monomers. Each monomer contains <scene name='HMG-CoA_Reductase/1dq8_star3_domains/2'>three domains </scene>, the <scene name='HMG-CoA_Reductase/1dq8_n_domain/2'>N-domain</scene>, the <scene name='HMG-CoA_Reductase/1dq8_l_domain/1'>L-Domain</scene>, and the <scene name='HMG-CoA_Reductase/1dq8_s_domain/1'>S-Domain</scene>. The L-domain is unique to HMGRs while the S-domain, which forms the binding site for NADP, resembles that of [[ferredoxin]]. The S and L domains are connected by a <scene name='HMG-CoA_Reductase/1dq8_cis_loop/6'>“cis-loop”</scene> which is essential for the HMG-binding site. <ref name="Roitelman"/> Salt bridges between residues R641 and E782 as well as <scene name='HMG-CoA_Reductase/1dq8_cis_loop/4'>hydrogen bonds</scene> between E700 and E700 on neighboring monomers compliment the largely hydrophobic dimer-dimer interface. <ref name="Roitelman"/> | ||
</StructureSection> | </StructureSection> | ||
| - | |||
| - | |||
| - | |||
| - | |||
<Structure load='1aoi' size='200' frame='true' align='right' caption='' /> | <Structure load='1aoi' size='200' frame='true' align='right' caption='' /> | ||
| Line 18: | Line 14: | ||
<tr> | <tr> | ||
<td> | <td> | ||
| - | <applet load='1aoi' size=' | + | <applet load='1aoi' size='300' frame='true' align='right' caption='' /> |
</td> | </td> | ||
<td> | <td> | ||
| - | <div style="background-color: #ffe8e8;overflow: auto;height: | + | <div style="background-color: #ffe8e8;overflow: auto;height: 300px;width: 100%;"> |
{{:User:Eran Hodis/Sandbox6}} | {{:User:Eran Hodis/Sandbox6}} | ||
</div> | </div> | ||
| Line 27: | Line 23: | ||
</tr> | </tr> | ||
</table> | </table> | ||
| + | |||
| + | ==References== | ||
| + | <references/> | ||
Revision as of 17:57, 31 October 2010
Contents |
Your Heading Here (maybe something like 'Structure')
| |||||||||||
|
Nucleosome Structure
|
IntroductionThe nucleosome core particle contains two copies of each histone protein (H2A, H2B, H3 and H4) and 146 basepairs (bp) of superhelical DNA wrapped around this histone octamer. It represents the first order of DNA packaging in the nucleus and as such is the principal structure that determines DNA accessibility. The Histone Octamer
alpha1-loop1-alpha2-loop2-alpha3 |
References
- ↑ Istvan ES, Deisenhofer J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science. 2001 May 11;292(5519):1160-4. PMID:11349148 doi:10.1126/science.1059344
- ↑ 2.0 2.1 2.2 2.3 Roitelman J, Olender EH, Bar-Nun S, Dunn WA Jr, Simoni RD. Immunological evidence for eight spans in the membrane domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase: implications for enzyme degradation in the endoplasmic reticulum. J Cell Biol. 1992 Jun;117(5):959-73. PMID:1374417
