This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
AndrewAlexanderSandbox1
From Proteopedia
(→Mechanism) |
|||
| Line 12: | Line 12: | ||
[[Image:Pyruvatekinasemechanism.gif|left]] | [[Image:Pyruvatekinasemechanism.gif|left]] | ||
<ref>{{website1| title=Exercise-Induced Metabolic Acidosis: Where do the Protons come from?|url=http://www.sportsci.org/jour/0102/rar.htm|}}</ref> | <ref>{{website1| title=Exercise-Induced Metabolic Acidosis: Where do the Protons come from?|url=http://www.sportsci.org/jour/0102/rar.htm|}}</ref> | ||
| - | <scene name='Keegan_Gelvoria_Sandbox_1/N_c_rainbow/null'>Pyruvate Kinase</scene> catalyzes the final reaction of glycolysis. It couples the free energy of PEP cleavage to the generation of ATP during the synthesis of the final product, pyruvate. | + | <scene name='Keegan_Gelvoria_Sandbox_1/N_c_rainbow/null'>Pyruvate Kinase</scene> catalyzes the final reaction of glycolysis. It couples the free energy of PEP cleavage to the generation of ATP during the synthesis of the final product, pyruvate.Pyruvate kinase cataylzes the transfer of phsphoryl group of PEP to ADP in the presence of two ions of MG2+ and one of K+. The first step is the nucleophilic attack of the PEP phosphorous atom by β-phosphoryl oxygen of ADP; this step displaces enolpyruvate while forming ATP. In the second step, enolpyruvate tautomerizes to pyruvate <ref>{{book |author=Voet, Donald; Voet, Judith C.; Pratt, Charlotte W.|title=Fundamentals of Biochemistry: Life at the Molecular Level|edition= 3|pages=502|}}</ref>. The formation of a high-energy intermediate by enolase in the 9th reaction of glycolysis allows for the synthesis of ATP in this reaction. Though the hydrolysis of 2PG is insufficient in driving the synthesis of ATP, the dehydration of 2PG allows for such a reaction to occur by forming a high-energy intermediate. The high potential of PEP reflects the large release of energy that occurs with the conversion of enolpyruvate to its keto tautomer, pyruvate <ref>{{book |author=Voet, Donald; Voet, Judith C.; Pratt, Charlotte W.|title=Fundamentals of Biochemistry: Life at the Molecular Level|edition= 3|pages=503|}}</ref>. |
==Kinetics and Regulation== | ==Kinetics and Regulation== | ||
| + | |||
| + | The kinetics of pyruvate kinase depend on K+ concnentration. <ref>{{article |author=Oria-Hernandez, Jesus.; Cabrera, Nallely.|title=Pyruvate KKinase Revisited: The Activating effect of K+|volume=280|pages=37924-37929|}}</ref>.: | ||
| + | Without a high K+ concnetration, the kinetic mechanism of pyruvate kinase changes from random to ordered with phosphoenolpyruvate as the first substrate. Vmax with K+ was about 400 times higher than a wild type sample without K+. In the presence of K+, the affinities for phosphoenolpruvate and ADP were 2-6 times higher than in the abscence of K+. This shows that K+ is involved in the acquisition of the active conformation of the enzyme, allowing either phosphoenolpyruvate or ADP to bind independently, but without K+, ADP cannot bind to the enzyme until phosphoenolpyruvate forms a competent active site for an ordered mechanism. Wild type pyruvate kinase without K+ has an ordered rapid equilibrium kinetic mechanism that shows Vmax to be 0.8 +/- -.04 umol/min mg with a kcat of 3.2 s^-1. When the wild-type pyruvate kinase has K+, it is in a random rapid equilibrium kinetic mechanism with a Vmax of 299 +/- 11 umol/min mg with a kcat of 1182. | ||
In the glycolytic cycle, there are three compounds that have a large negative ∆G which includes the reaction pyruvate kinase catalyzes. Due to these three steps regulating the overall activity of the cycle, they are generally irreversible in vivo. Through numerous studies, the activity of pyruvate kinase has been found to be regulated by these effectors <ref>{{article |author=Dann, Leighton G.; Britton, Hubert G.|title=Kinetics and Mechanism of Action of Muscle Pyruvate Kinase|volume=169|pages=39-54|}}</ref>.: | In the glycolytic cycle, there are three compounds that have a large negative ∆G which includes the reaction pyruvate kinase catalyzes. Due to these three steps regulating the overall activity of the cycle, they are generally irreversible in vivo. Through numerous studies, the activity of pyruvate kinase has been found to be regulated by these effectors <ref>{{article |author=Dann, Leighton G.; Britton, Hubert G.|title=Kinetics and Mechanism of Action of Muscle Pyruvate Kinase|volume=169|pages=39-54|}}</ref>.: | ||
| Line 27: | Line 30: | ||
| - | As indicated earlier, phosphoenolpyruvate can enhance the activity of the reaction by adding into the enzyme because it is the rate limiting step. The enzyme follows hyperbolic kinetics. Experiments found that no incorporation was found in the reaction, indicating a random, rapid dissociation of the products. This, then, assumes that the products inhibit the enzyme’s reaction by simply reversing the reaction. Both pyruvate and ATP have been shown to be non-competitive inhibitors of pyruvate kinase. <ref>{{article |author=Dann, Leighton G.; Britton, Hubert G.|title=Kinetics and Mechanism of Action of Muscle Pyruvate Kinase|volume=169|pages=39-54|}}</ref>. | + | As indicated earlier, phosphoenolpyruvate can enhance the activity of the reaction by adding into the enzyme because it is the rate limiting step. The enzyme follows hyperbolic kinetics. Experiments found that no incorporation was found in the reaction, indicating a random, rapid dissociation of the products. This, then, assumes that the products inhibit the enzyme’s reaction by simply reversing the reaction. Both pyruvate and ATP have been shown to be non-competitive inhibitors of pyruvate kinase. <ref>{{article |author=Dann, Leighton G.; Britton, Hubert G.|title=Kinetics and Mechanism of Action of Muscle Pyruvate Kinase|volume=169|pages=39-54|}}</ref>. With or without K+, oxalate is a noncompetitive inhibitor of ADP-Mg2+. It will form a nonproductive ternary complex which diminishes Vmax without altering the binding of ADP-Mg2+.ref>{{article |author=Oria-Hernandez, Jesus.; Cabrera, Nallely.|title=Pyruvate KKinase Revisited: The Activating effect of K+|volume=280|pages=37924-37929|}}</ref>.: |
| + | |||
==Pyruvate Kinase Deficiency== | ==Pyruvate Kinase Deficiency== | ||
Revision as of 17:58, 17 April 2011
Contents |
Pyruvate Kinase
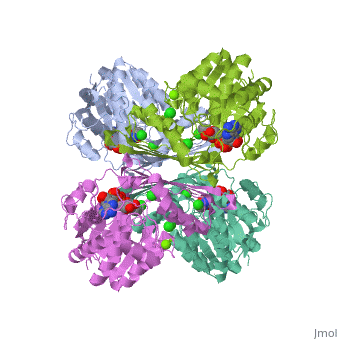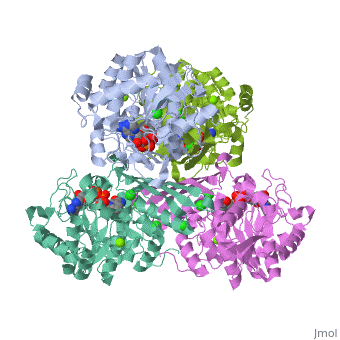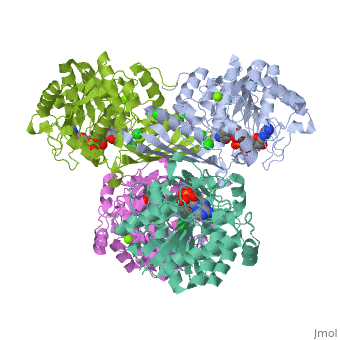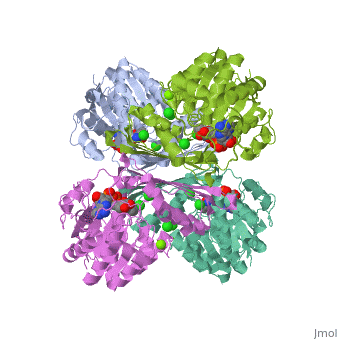
Pyruvate Kinase is an enzyme that is involved in glycolysis. Pyruvate kinase’s function is to catalyze the last step of glycolysis; thereby, generating the second ATP of glycolysis and pyruvate. It is able to catalyze this step by transferring the phosphate group from phosphoenolpyruvate (PEP) to ADP [1].
| |||||||||
| 2vgb, resolution 2.73Å () | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ligands: | , , , | ||||||||
| Activity: | Pyruvate kinase, with EC number 2.7.1.40 | ||||||||
| Related: | 1liy, 1liu, 1liw, 1lix | ||||||||
| |||||||||
| |||||||||
| Resources: | FirstGlance, OCA, RCSB, PDBsum | ||||||||
| Coordinates: | save as pdb, mmCIF, xml | ||||||||
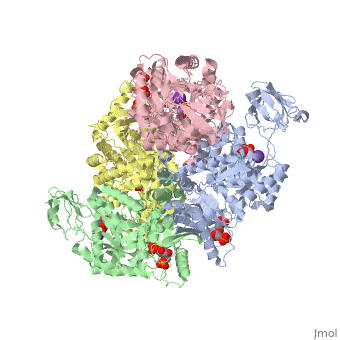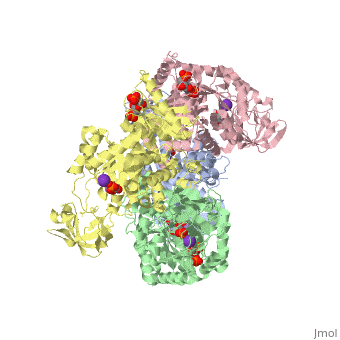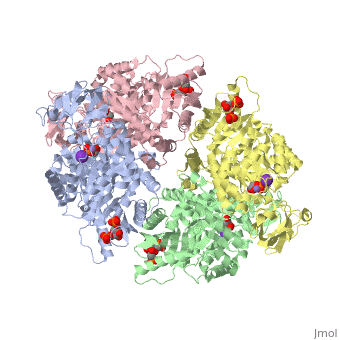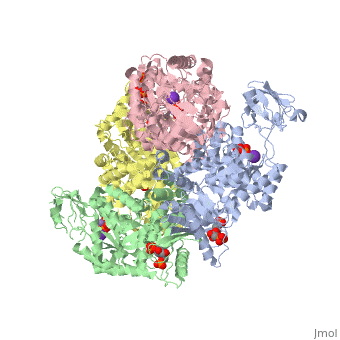

Structure
This particular protein is found in Homo sapiens and has the abbreviation PK. Pyruvate kinase belongs to the all beta proteins class and has the PK beta-barrel domain-like fold. It belongs to the PK beta-barrel domain-like superfamily and pyruvate kinase beta-barrel domain family[2].
Though pyruvate kinase is classified into all beta proteins, pyruvate kinase's comprises of both alpha helices and beta sheets. In the quaternary structure of pyruvate kinase, it can be observed to have in humans. Thus, this enzyme is tetrameric with on each domain for the and Mg2+ ligands to bind to. There are four types of tissue-specific isozymes: L (liver), R (red blood cells), M1 (muscle, heart, and brain), and M2 (early fetal tissue)[3].
Mechanism

[4] catalyzes the final reaction of glycolysis. It couples the free energy of PEP cleavage to the generation of ATP during the synthesis of the final product, pyruvate.Pyruvate kinase cataylzes the transfer of phsphoryl group of PEP to ADP in the presence of two ions of MG2+ and one of K+. The first step is the nucleophilic attack of the PEP phosphorous atom by β-phosphoryl oxygen of ADP; this step displaces enolpyruvate while forming ATP. In the second step, enolpyruvate tautomerizes to pyruvate [5]. The formation of a high-energy intermediate by enolase in the 9th reaction of glycolysis allows for the synthesis of ATP in this reaction. Though the hydrolysis of 2PG is insufficient in driving the synthesis of ATP, the dehydration of 2PG allows for such a reaction to occur by forming a high-energy intermediate. The high potential of PEP reflects the large release of energy that occurs with the conversion of enolpyruvate to its keto tautomer, pyruvate [6].
Kinetics and Regulation
The kinetics of pyruvate kinase depend on K+ concnentration. [7].:
Without a high K+ concnetration, the kinetic mechanism of pyruvate kinase changes from random to ordered with phosphoenolpyruvate as the first substrate. Vmax with K+ was about 400 times higher than a wild type sample without K+. In the presence of K+, the affinities for phosphoenolpruvate and ADP were 2-6 times higher than in the abscence of K+. This shows that K+ is involved in the acquisition of the active conformation of the enzyme, allowing either phosphoenolpyruvate or ADP to bind independently, but without K+, ADP cannot bind to the enzyme until phosphoenolpyruvate forms a competent active site for an ordered mechanism. Wild type pyruvate kinase without K+ has an ordered rapid equilibrium kinetic mechanism that shows Vmax to be 0.8 +/- -.04 umol/min mg with a kcat of 3.2 s^-1. When the wild-type pyruvate kinase has K+, it is in a random rapid equilibrium kinetic mechanism with a Vmax of 299 +/- 11 umol/min mg with a kcat of 1182.
In the glycolytic cycle, there are three compounds that have a large negative ∆G which includes the reaction pyruvate kinase catalyzes. Due to these three steps regulating the overall activity of the cycle, they are generally irreversible in vivo. Through numerous studies, the activity of pyruvate kinase has been found to be regulated by these effectors [8].:
a. Phosphoenolpyruvate, the substrate, can impact enzymatic activity by enhancing the reaction by allowing the process to operate faster with more substrate present.
b. ATP and pyruvate have been found to be a negative allosteric inhibitor.
c. Alanine has also been found to be a negative allosteric modulator.
As indicated earlier, phosphoenolpyruvate can enhance the activity of the reaction by adding into the enzyme because it is the rate limiting step. The enzyme follows hyperbolic kinetics. Experiments found that no incorporation was found in the reaction, indicating a random, rapid dissociation of the products. This, then, assumes that the products inhibit the enzyme’s reaction by simply reversing the reaction. Both pyruvate and ATP have been shown to be non-competitive inhibitors of pyruvate kinase. [9]. With or without K+, oxalate is a noncompetitive inhibitor of ADP-Mg2+. It will form a nonproductive ternary complex which diminishes Vmax without altering the binding of ADP-Mg2+.ref>Dann, Leighton G. and Britton, Hubert G. Kinetics and Mechanism of Action of Muscle Pyruvate Kinase. Journal of Biochemistry. 1978 169:39-54.</ref>.:
Pyruvate Kinase Deficiency
Pyruvate kinase deficiency is the most frequent enzyme abnormality of glycolysis that causes hemolytic anemia. In cells that lack mitochondria, this deficiency is especially harmful, because these cells must use anaerobic glycolysis as their sole source of energy because the TCA cycle is not available. Red blood cells, in a state of pyruvate kinase deficiency, rapidly become deficient in ATP and can undergo hemolysis.This is transmitted as an autosomal recessive trit. The severity of haemolysis is extremely variable such as a mild case to life-threatening neonatal anaemia requiring transfusions. Over one hundred eighty different mutations have been discovered in relation to this deficiency with most being autosomal recessive, but a few strands are autosomal dominant. The deficiency causes red blood cells to deform into echinocytes on peripheral blood smears. This causes the buildup of reaction intermediates which can also increase the level of 2,3-bisphosphoglycerate in the cells. This causes a rightward shift in the hemoglobin oxygen saturation curve, which means that there is a decreased oxygen affinity for the hemoglobin and earlier oxygen unloading than under normal conditions.
[10].
Additional Resources
For additional information, see: Carbohydrate Metabolism
References
- ↑ Voet, Donald, Judith G. Voet, and Charlotte W. Pratt. Fundamentals of Biochemistry: Life at the Molecular Level. 3rd ed. Hoboken, NJ: John Wiley & Sons, Inc., 2008, 501-503.
- ↑ authors, The scop. "Structural Classification of Proteins". 2009. 2/26 2010. <http://scop.berkeley.edu/data/scop.b.c.jh.b.b.d.html>.
- ↑ authors, The scop. "Structural Classification of Proteins". 2009. 2/26 2010. <http://scop.berkeley.edu/data/scop.b.c.jh.b.b.d.html>.
- ↑ Robergs, Robert. "Exercise-Induced Metabolic Acidosis: Where do the Protons come from?". 2009. 2/27 2010. <http://www.sportsci.org/jour/0102/rar.htm>.
- ↑ Voet, Donald, Judith G. Voet, and Charlotte W. Pratt. Fundamentals of Biochemistry: Life at the Molecular Level. 3rd ed. Hoboken, NJ: John Wiley & Sons, Inc., 2008, 501-503.
- ↑ Voet, Donald, Judith G. Voet, and Charlotte W. Pratt. Fundamentals of Biochemistry: Life at the Molecular Level. 3rd ed. Hoboken, NJ: John Wiley & Sons, Inc., 2008, 501-503.
- ↑ Dann, Leighton G. and Britton, Hubert G. Kinetics and Mechanism of Action of Muscle Pyruvate Kinase. Journal of Biochemistry. 1978 169:39-54.
- ↑ Dann, Leighton G. and Britton, Hubert G. Kinetics and Mechanism of Action of Muscle Pyruvate Kinase. Journal of Biochemistry. 1978 169:39-54.
- ↑ Dann, Leighton G. and Britton, Hubert G. Kinetics and Mechanism of Action of Muscle Pyruvate Kinase. Journal of Biochemistry. 1978 169:39-54.
- ↑ Dann, Leighton G. and Britton, Hubert G. Kinetics and Mechanism of Action of Muscle Pyruvate Kinase. Journal of Biochemistry. 1978 169:39-54.
Replace the PDB id (use lowercase!) after the STRUCTURE_ and after PDB= to load and display another structure.
| |||||||||
| 3cin, resolution 1.70Å () | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ligands: | , , | ||||||||
| Gene: | TM1419, TM_1419 (Thermotoga maritima MSB8) | ||||||||
| Activity: | Inositol-3-phosphate synthase, with EC number 5.5.1.4 | ||||||||
| |||||||||
| |||||||||
| Resources: | FirstGlance, OCA, RCSB, PDBsum, TOPSAN | ||||||||
| Coordinates: | save as pdb, mmCIF, xml | ||||||||