This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
ToxT
From Proteopedia
| Line 9: | Line 9: | ||
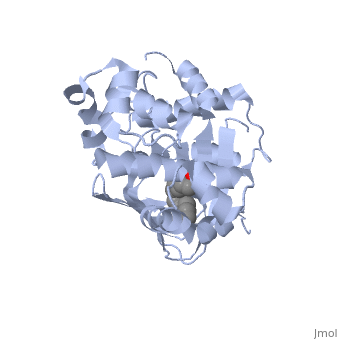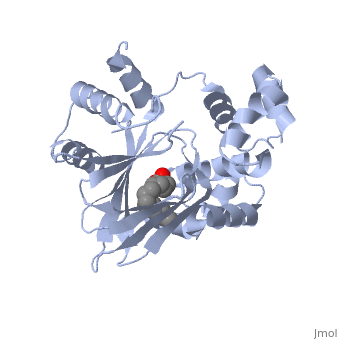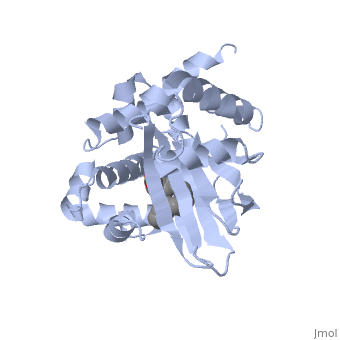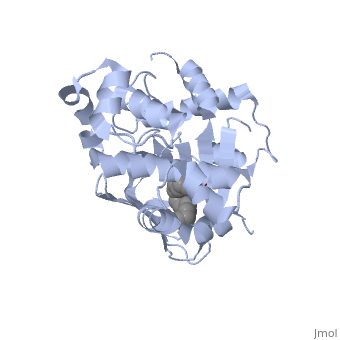
<u>Ligand-binding</u>. <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Barrel/1'>A nine-stranded beta sheet sandwich</scene> or "jelly-roll" with three other alpha helices (overall making up the <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/N-terminus/1'>N-terminus</scene>) contain a <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Binding_pocket/1'>binding pocket</scene>. This is made from several residues from the N-terminus (Y12, Y20, F22, L25, I27, K31, F33, L61, F69, L71, V81, and V83), and a few from the C-terminus (I226, K230, M259, V261, Y266, and M269). This pocket contains a sixteen-carbon fatty acid positioned in a conformation such that its negatively charged carboxylate group forms salt bridges between K31 of the N-terminal domain, and K230 from the C-terminal domain. The pocket is highly hydrophobic, and has a known volume of 780.9 Angstroms.<ref name="structure">PMID: 20133655</ref> This pocket contains a ligand: <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Binding_pocket/2'>cis-palmitoleate</scene> <ref name="structure">PMID: 20133655</ref> which appears to have a negative effect on virulence when present in vitro. The <i>cis</i>-palmitoleate forms <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Salt_bridges_pam/1'>salt bridges</scene> with residues K31 and K230 (for detail, see Figure 1B of: [http://www.pnas.org/content/107/7/2860/F1.large.jpg]). This unsaturated fatty acid, like other UFAs,[http://en.wikipedia.org/wiki/Fatty_acid#Unsaturated_fatty_acids] tend to inhibit genes under the control of ToxT. | <u>Ligand-binding</u>. <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Barrel/1'>A nine-stranded beta sheet sandwich</scene> or "jelly-roll" with three other alpha helices (overall making up the <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/N-terminus/1'>N-terminus</scene>) contain a <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Binding_pocket/1'>binding pocket</scene>. This is made from several residues from the N-terminus (Y12, Y20, F22, L25, I27, K31, F33, L61, F69, L71, V81, and V83), and a few from the C-terminus (I226, K230, M259, V261, Y266, and M269). This pocket contains a sixteen-carbon fatty acid positioned in a conformation such that its negatively charged carboxylate group forms salt bridges between K31 of the N-terminal domain, and K230 from the C-terminal domain. The pocket is highly hydrophobic, and has a known volume of 780.9 Angstroms.<ref name="structure">PMID: 20133655</ref> This pocket contains a ligand: <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Binding_pocket/2'>cis-palmitoleate</scene> <ref name="structure">PMID: 20133655</ref> which appears to have a negative effect on virulence when present in vitro. The <i>cis</i>-palmitoleate forms <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Salt_bridges_pam/1'>salt bridges</scene> with residues K31 and K230 (for detail, see Figure 1B of: [http://www.pnas.org/content/107/7/2860/F1.large.jpg]). This unsaturated fatty acid, like other UFAs,[http://en.wikipedia.org/wiki/Fatty_acid#Unsaturated_fatty_acids] tend to inhibit genes under the control of ToxT. | ||
Specifically, the <i>cis</i>-palmitoleate (PAM) appears to change ToxT's conformation, and thus lower its ability to bind DNA and form dimers.<ref name="structure">PMID: 20133655</ref> The presence of UFAs is associated with being in the lumen of the intestine during the bacterial infection. PAM brings K31 and K230 together from either end of the protein, and essentially closes off ToxT. K230 is at the end of helix seven, and binding to K31 causes helix six to be pulled into an unfavorable conformation that deters DNA binding. In lower concentration of fatty acids, ie: after penetrating the intestine's mucus, PAM is in lower concentration. At this point, charge-charge repulsion between K31 and K230 leads to a destabilization of the closed conformation of ToxT. This repulsion prompts the opening of the N and C terminal domains. The freedom of helices six and seven to find a favorable configuration allows DNA binding to occur.<ref name="structure">PMID: 20133655</ref> | Specifically, the <i>cis</i>-palmitoleate (PAM) appears to change ToxT's conformation, and thus lower its ability to bind DNA and form dimers.<ref name="structure">PMID: 20133655</ref> The presence of UFAs is associated with being in the lumen of the intestine during the bacterial infection. PAM brings K31 and K230 together from either end of the protein, and essentially closes off ToxT. K230 is at the end of helix seven, and binding to K31 causes helix six to be pulled into an unfavorable conformation that deters DNA binding. In lower concentration of fatty acids, ie: after penetrating the intestine's mucus, PAM is in lower concentration. At this point, charge-charge repulsion between K31 and K230 leads to a destabilization of the closed conformation of ToxT. This repulsion prompts the opening of the N and C terminal domains. The freedom of helices six and seven to find a favorable configuration allows DNA binding to occur.<ref name="structure">PMID: 20133655</ref> | ||
| - | <br/> | + | <br/> [[Image:MSA.png]] |
<br/> | <br/> | ||
<u>Dimerization</u>. Though the structure shown is a monomer with two overall domains (N-terminal and C-terminal), ToxT tends to form a dimer.<ref name="dimerization">PMID: 21415495 | <u>Dimerization</u>. Though the structure shown is a monomer with two overall domains (N-terminal and C-terminal), ToxT tends to form a dimer.<ref name="dimerization">PMID: 21415495 | ||
</ref> The preferred state of ToxT varies between promoters, but binding to the <i>ctx</i> promoter to generate cholera toxin appears to be possible only in the dimer form.<ref name="virstatin">PMID:17283330</ref>ToxT binds to thirteen base pair sequences (can be single, direct, or inverted repeats) called toxboxes in order to activate their respective promoters.[http://www.sigwiki.info/wiki/Signature:ToxBox] | </ref> The preferred state of ToxT varies between promoters, but binding to the <i>ctx</i> promoter to generate cholera toxin appears to be possible only in the dimer form.<ref name="virstatin">PMID:17283330</ref>ToxT binds to thirteen base pair sequences (can be single, direct, or inverted repeats) called toxboxes in order to activate their respective promoters.[http://www.sigwiki.info/wiki/Signature:ToxBox] | ||
==Ligand== | ==Ligand== | ||
| - | In this resolved structure, <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Pam/2'>cis-palmitoleate</scene>[http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=3gbg&template=ligands.html&l=1.1] is shown, which can be bound in the beta sheet barrel (as discussed above). This unsaturated fatty acid reduces virulence expression in <i>Vibrio cholerae</i>. | + | In this resolved structure, <scene name='ToxT_Transcriptional_Regulator_in_Vibrio_cholerae/Pam/2'>cis-palmitoleate</scene>[http://www.ebi.ac.uk/thornton-srv/databases/cgi-bin/pdbsum/GetPage.pl?pdbcode=3gbg&template=ligands.html&l=1.1] is shown, which can be bound in the beta sheet barrel (as discussed above). This unsaturated fatty acid reduces virulence expression in <i>Vibrio cholerae</i>. |
</StructureSection> | </StructureSection> | ||
==Further Study== | ==Further Study== | ||
Revision as of 06:14, 5 May 2012
The crystal structure of ToxT is resolved in monomeric form, after isolation from Vibrio cholerae strain O395.[1]
Introduction
ToxT is a molecule at the end of a transcriptional cascade that autoregulates the transcription of the primary virulence factors of Vibrio cholerae[1] and itself. ToxT is a cytoplasmic protein that is activated in turn by ToxR, which is itself activated by ToxS in response to environmental stimuli.[2] These two factors, cholera toxin (CT)[2] and the toxin co-regulated pilus (TCP), are instrumental in causing the disease cholera[3]. This is an intestinal infection resulting in massive water loss in the affected individual, causing extreme dehydration.[4] Rehydration is sufficient as treatment.
| |||||||||||


Further Study
Conclusive results about what activates ToxT itself has not yet been found. The varying activity of ToxT dependent on the presence of cis-palmitoleate or other unsaturated fatty acids represents a detailed method of effective pathogenicity in humans, but may not be a reasonable target for drug treatment. By restricting transcription (and thus translation and protein production) of virulence genes until the bacterium is determined to be in a favorable location for infection, Vibrio cholerae avoids wasting energy producing virulence factors that will just be cleared by the intestine. This is a specific mechanism to ensure that the bacterium also injects CT and TCP where they will do the most damage, perpetuating the infection. [6] Despite the lack of information about what activates ToxT itself, it is understood that the transcription of ctxA and tcpA by vibrio cholerae is sharply reduced in the presence of oleic, linoleic acid, and arachidonic acid, all of which are components of bile. Therefore, one may hypothesize that it may be possible to use the structure of a UFA or SFA to design a small molecule inhibitor of ToxT which may be used to treat or prevent cholera.
Evolution
Vibrio cholerae is a highly diverse species in which some strains are completely harmless, whereas other strains have the capacity to cause global cholera pandemics. It has been discovered that in each epidemic and pandemic strain, there is a chromosomal pathogenicity island (PAI) that is not present in the nonpathogenic strains. The region containing two ToxR-regulated genes (aldA and tagA) is composed of 13kb of previously unidentified DNA. This region is part of a PAI that contains ToxT and a gene cluster a critical colonization factor and TCP. The PAI is 39.5 kb long, contains putative integrase and transposase genes, and inserts near a 10Sa RNA gene. One may infer that the PAI could have originated from a bacteriophage. This PAI was also found in two non-O1/non-O139 (which are both pandemic) sero type strains. Therefore, one may hypothesize that the PAI could be transferred within other strains of Vibrio cholerae.

Proteopedia Page Contributors and Editors (what is this?)
Ingrid Youngworth, Yang Yang, Michal Harel, Alexander Berchansky, Jaime Prilusky

![[1]](http://www.pnas.org/content/107/7/2860/F3.large.jpg){kind=link}
![[2]](http://www.pnas.org/content/107/7/2860/F1.large.jpg){kind=link}