1q21
From Proteopedia
| Line 1: | Line 1: | ||
| - | [[Image:1q21.png|left|200px]] | ||
| - | |||
{{STRUCTURE_1q21| PDB=1q21 | SCENE= }} | {{STRUCTURE_1q21| PDB=1q21 | SCENE= }} | ||
| - | |||
===CRYSTAL STRUCTURES AT 2.2 ANGSTROMS RESOLUTION OF THE CATALYTIC DOMAINS OF NORMAL RAS PROTEIN AND AN ONCOGENIC MUTANT COMPLEXED WITH GSP=== | ===CRYSTAL STRUCTURES AT 2.2 ANGSTROMS RESOLUTION OF THE CATALYTIC DOMAINS OF NORMAL RAS PROTEIN AND AN ONCOGENIC MUTANT COMPLEXED WITH GSP=== | ||
| + | {{ABSTRACT_PUBMED_1899707}} | ||
| - | + | ==Disease== | |
| + | [[http://www.uniprot.org/uniprot/RASH_HUMAN RASH_HUMAN]] Defects in HRAS are the cause of faciocutaneoskeletal syndrome (FCSS) [MIM:[http://omim.org/entry/218040 218040]]. A rare condition characterized by prenatally increased growth, postnatal growth deficiency, mental retardation, distinctive facial appearance, cardiovascular abnormalities (typically pulmonic stenosis, hypertrophic cardiomyopathy and/or atrial tachycardia), tumor predisposition, skin and musculoskeletal abnormalities.<ref>PMID:16170316</ref><ref>PMID:16329078</ref><ref>PMID:16443854</ref><ref>PMID:17054105</ref><ref>PMID:18247425</ref><ref>PMID:18039947</ref><ref>PMID:19995790</ref> Defects in HRAS are the cause of congenital myopathy with excess of muscle spindles (CMEMS) [MIM:[http://omim.org/entry/218040 218040]]. CMEMS is a variant of Costello syndrome.<ref>PMID:17412879</ref> Defects in HRAS may be a cause of susceptibility to Hurthle cell thyroid carcinoma (HCTC) [MIM:[http://omim.org/entry/607464 607464]]. Hurthle cell thyroid carcinoma accounts for approximately 3% of all thyroid cancers. Although they are classified as variants of follicular neoplasms, they are more often multifocal and somewhat more aggressive and are less likely to take up iodine than are other follicular neoplasms. Note=Mutations which change positions 12, 13 or 61 activate the potential of HRAS to transform cultured cells and are implicated in a variety of human tumors. Defects in HRAS are a cause of susceptibility to bladder cancer (BLC) [MIM:[http://omim.org/entry/109800 109800]]. A malignancy originating in tissues of the urinary bladder. It often presents with multiple tumors appearing at different times and at different sites in the bladder. Most bladder cancers are transitional cell carcinomas. They begin in cells that normally make up the inner lining of the bladder. Other types of bladder cancer include squamous cell carcinoma (cancer that begins in thin, flat cells) and adenocarcinoma (cancer that begins in cells that make and release mucus and other fluids). Bladder cancer is a complex disorder with both genetic and environmental influences. Note=Defects in HRAS are the cause of oral squamous cell carcinoma (OSCC).<ref>PMID:1459726</ref> Defects in HRAS are the cause of Schimmelpenning-Feuerstein-Mims syndrome (SFM) [MIM:[http://omim.org/entry/163200 163200]]. A disease characterized by sebaceous nevi, often on the face, associated with variable ipsilateral abnormalities of the central nervous system, ocular anomalies, and skeletal defects. Many oral manifestations have been reported, not only including hypoplastic and malformed teeth, and mucosal papillomatosis, but also ankyloglossia, hemihyperplastic tongue, intraoral nevus, giant cell granuloma, ameloblastoma, bone cysts, follicular cysts, oligodontia, and odontodysplasia. Sebaceous nevi follow the lines of Blaschko and these can continue as linear intraoral lesions, as in mucosal papillomatosis.<ref>PMID:22683711</ref> | ||
| + | |||
| + | ==Function== | ||
| + | [[http://www.uniprot.org/uniprot/RASH_HUMAN RASH_HUMAN]] Ras proteins bind GDP/GTP and possess intrinsic GTPase activity.<ref>PMID:14500341</ref><ref>PMID:9020151</ref><ref>PMID:12740440</ref> | ||
==About this Structure== | ==About this Structure== | ||
| Line 14: | Line 16: | ||
==Reference== | ==Reference== | ||
| - | <ref group="xtra">PMID:001899707</ref><ref group="xtra">PMID:009398520</ref><ref group="xtra">PMID:009428770</ref><references group="xtra"/> | + | <ref group="xtra">PMID:001899707</ref><ref group="xtra">PMID:009398520</ref><ref group="xtra">PMID:009428770</ref><references group="xtra"/><references/> |
[[Category: Homo sapiens]] | [[Category: Homo sapiens]] | ||
[[Category: Kim, S H.]] | [[Category: Kim, S H.]] | ||
[[Category: Oncogene protein]] | [[Category: Oncogene protein]] | ||
Revision as of 18:11, 24 March 2013
| |||||||||
| 1q21, resolution 2.20Å () | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ligands: | , | ||||||||
| |||||||||
| |||||||||
| Resources: | FirstGlance, OCA, RCSB, PDBsum | ||||||||
| Coordinates: | save as pdb, mmCIF, xml | ||||||||
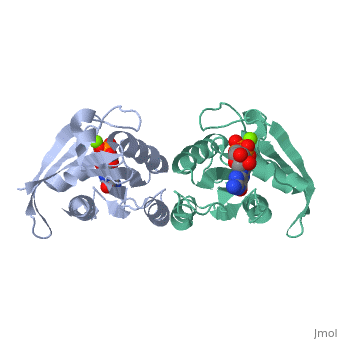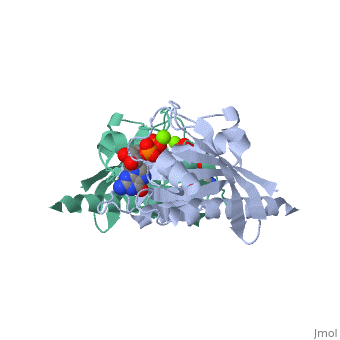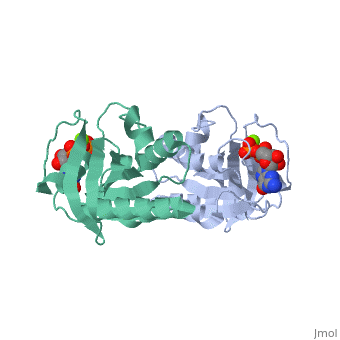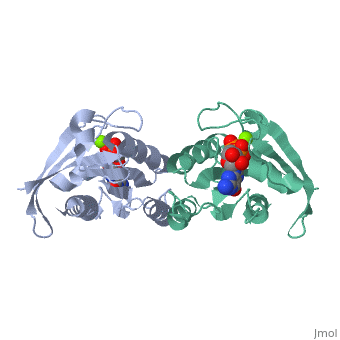

Contents |
CRYSTAL STRUCTURES AT 2.2 ANGSTROMS RESOLUTION OF THE CATALYTIC DOMAINS OF NORMAL RAS PROTEIN AND AN ONCOGENIC MUTANT COMPLEXED WITH GSP
The biological functions of ras proteins are controlled by the bound guanine nucleotide GDP or GTP. The GTP-bound conformation is biologically active, and is rapidly deactivated to the GDP-bound conformation through interaction with GAP (GTPase Activating Protein). Most transforming mutants of ras proteins have drastically reduced GTP hydrolysis rates even in the presence of GAP. The crystal structures of the GDP complexes of ras proteins at 2.2 A resolution reveal the detailed interaction between the ras proteins and the GDP molecule. All the currently known transforming mutation positions are clustered around the bound guanine nucleotide molecule. The presumed "effector" region and the GAP recognition region are both highly exposed. No significant structural differences were found between the GDP complexes of normal ras protein and the oncogenic mutant with valine at position 12, except the side-chain of the valine residue. However, comparison with GTP-analog complexes of ras proteins suggests that the valine side-chain may inhibit GTP hydrolysis in two possible ways: (1) interacting directly with the gamma-phosphate and altering its orientation or the conformation of protein residues around the phosphates; and/or (2) preventing either the departure of gamma-phosphate on GTP hydrolysis or the entrance of a nucleophilic group to attack the gamma-phosphate. The structural similarity between ras protein and the bacterial elongation factor Tu suggests that their common structural motif might be conserved for other guanine nucleotide binding proteins.
Crystal structures at 2.2 A resolution of the catalytic domains of normal ras protein and an oncogenic mutant complexed with GDP., Tong LA, de Vos AM, Milburn MV, Kim SH, J Mol Biol. 1991 Feb 5;217(3):503-16. PMID:1899707
From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine.
Disease
[RASH_HUMAN] Defects in HRAS are the cause of faciocutaneoskeletal syndrome (FCSS) [MIM:218040]. A rare condition characterized by prenatally increased growth, postnatal growth deficiency, mental retardation, distinctive facial appearance, cardiovascular abnormalities (typically pulmonic stenosis, hypertrophic cardiomyopathy and/or atrial tachycardia), tumor predisposition, skin and musculoskeletal abnormalities.[1][2][3][4][5][6][7] Defects in HRAS are the cause of congenital myopathy with excess of muscle spindles (CMEMS) [MIM:218040]. CMEMS is a variant of Costello syndrome.[8] Defects in HRAS may be a cause of susceptibility to Hurthle cell thyroid carcinoma (HCTC) [MIM:607464]. Hurthle cell thyroid carcinoma accounts for approximately 3% of all thyroid cancers. Although they are classified as variants of follicular neoplasms, they are more often multifocal and somewhat more aggressive and are less likely to take up iodine than are other follicular neoplasms. Note=Mutations which change positions 12, 13 or 61 activate the potential of HRAS to transform cultured cells and are implicated in a variety of human tumors. Defects in HRAS are a cause of susceptibility to bladder cancer (BLC) [MIM:109800]. A malignancy originating in tissues of the urinary bladder. It often presents with multiple tumors appearing at different times and at different sites in the bladder. Most bladder cancers are transitional cell carcinomas. They begin in cells that normally make up the inner lining of the bladder. Other types of bladder cancer include squamous cell carcinoma (cancer that begins in thin, flat cells) and adenocarcinoma (cancer that begins in cells that make and release mucus and other fluids). Bladder cancer is a complex disorder with both genetic and environmental influences. Note=Defects in HRAS are the cause of oral squamous cell carcinoma (OSCC).[9] Defects in HRAS are the cause of Schimmelpenning-Feuerstein-Mims syndrome (SFM) [MIM:163200]. A disease characterized by sebaceous nevi, often on the face, associated with variable ipsilateral abnormalities of the central nervous system, ocular anomalies, and skeletal defects. Many oral manifestations have been reported, not only including hypoplastic and malformed teeth, and mucosal papillomatosis, but also ankyloglossia, hemihyperplastic tongue, intraoral nevus, giant cell granuloma, ameloblastoma, bone cysts, follicular cysts, oligodontia, and odontodysplasia. Sebaceous nevi follow the lines of Blaschko and these can continue as linear intraoral lesions, as in mucosal papillomatosis.[10]
Function
[RASH_HUMAN] Ras proteins bind GDP/GTP and possess intrinsic GTPase activity.[11][12][13]
About this Structure
1q21 is a 1 chain structure with sequence from Homo sapiens. This structure supersedes the now removed PDB entry 2p21. Full crystallographic information is available from OCA.
See Also
Reference
- Tong LA, de Vos AM, Milburn MV, Kim SH. Crystal structures at 2.2 A resolution of the catalytic domains of normal ras protein and an oncogenic mutant complexed with GDP. J Mol Biol. 1991 Feb 5;217(3):503-16. PMID:1899707
- Ma J, Karplus M. Ligand-induced conformational changes in ras p21: a normal mode and energy minimization analysis. J Mol Biol. 1997 Nov 21;274(1):114-31. PMID:9398520 doi:10.1006/jmbi.1997.1313
- Lowe J, Amos LA. Crystal structure of the bacterial cell-division protein FtsZ. Nature. 1998 Jan 8;391(6663):203-6. PMID:9428770 doi:10.1038/34472
- ↑ Aoki Y, Niihori T, Kawame H, Kurosawa K, Ohashi H, Tanaka Y, Filocamo M, Kato K, Suzuki Y, Kure S, Matsubara Y. Germline mutations in HRAS proto-oncogene cause Costello syndrome. Nat Genet. 2005 Oct;37(10):1038-40. Epub 2005 Sep 18. PMID:16170316 doi:ng1641
- ↑ Gripp KW, Lin AE, Stabley DL, Nicholson L, Scott CI Jr, Doyle D, Aoki Y, Matsubara Y, Zackai EH, Lapunzina P, Gonzalez-Meneses A, Holbrook J, Agresta CA, Gonzalez IL, Sol-Church K. HRAS mutation analysis in Costello syndrome: genotype and phenotype correlation. Am J Med Genet A. 2006 Jan 1;140(1):1-7. PMID:16329078 doi:10.1002/ajmg.a.31047
- ↑ Kerr B, Delrue MA, Sigaudy S, Perveen R, Marche M, Burgelin I, Stef M, Tang B, Eden OB, O'Sullivan J, De Sandre-Giovannoli A, Reardon W, Brewer C, Bennett C, Quarell O, M'Cann E, Donnai D, Stewart F, Hennekam R, Cave H, Verloes A, Philip N, Lacombe D, Levy N, Arveiler B, Black G. Genotype-phenotype correlation in Costello syndrome: HRAS mutation analysis in 43 cases. J Med Genet. 2006 May;43(5):401-5. Epub 2006 Jan 27. PMID:16443854 doi:jmg.2005.040352
- ↑ Zampino G, Pantaleoni F, Carta C, Cobellis G, Vasta I, Neri C, Pogna EA, De Feo E, Delogu A, Sarkozy A, Atzeri F, Selicorni A, Rauen KA, Cytrynbaum CS, Weksberg R, Dallapiccola B, Ballabio A, Gelb BD, Neri G, Tartaglia M. Diversity, parental germline origin, and phenotypic spectrum of de novo HRAS missense changes in Costello syndrome. Hum Mutat. 2007 Mar;28(3):265-72. PMID:17054105 doi:10.1002/humu.20431
- ↑ Gripp KW, Innes AM, Axelrad ME, Gillan TL, Parboosingh JS, Davies C, Leonard NJ, Lapointe M, Doyle D, Catalano S, Nicholson L, Stabley DL, Sol-Church K. Costello syndrome associated with novel germline HRAS mutations: an attenuated phenotype? Am J Med Genet A. 2008 Mar 15;146A(6):683-90. PMID:18247425 doi:10.1002/ajmg.a.32227
- ↑ Lo IF, Brewer C, Shannon N, Shorto J, Tang B, Black G, Soo MT, Ng DK, Lam ST, Kerr B. Severe neonatal manifestations of Costello syndrome. J Med Genet. 2008 Mar;45(3):167-71. Epub 2007 Nov 26. PMID:18039947 doi:10.1136/jmg.2007.054411
- ↑ Gremer L, De Luca A, Merbitz-Zahradnik T, Dallapiccola B, Morlot S, Tartaglia M, Kutsche K, Ahmadian MR, Rosenberger G. Duplication of Glu37 in the switch I region of HRAS impairs effector/GAP binding and underlies Costello syndrome by promoting enhanced growth factor-dependent MAPK and AKT activation. Hum Mol Genet. 2010 Mar 1;19(5):790-802. doi: 10.1093/hmg/ddp548. Epub 2009 Dec, 8. PMID:19995790 doi:10.1093/hmg/ddp548
- ↑ van der Burgt I, Kupsky W, Stassou S, Nadroo A, Barroso C, Diem A, Kratz CP, Dvorsky R, Ahmadian MR, Zenker M. Myopathy caused by HRAS germline mutations: implications for disturbed myogenic differentiation in the presence of constitutive HRas activation. J Med Genet. 2007 Jul;44(7):459-62. Epub 2007 Apr 5. PMID:17412879 doi:jmg.2007.049270
- ↑ Sakai E, Rikimaru K, Ueda M, Matsumoto Y, Ishii N, Enomoto S, Yamamoto H, Tsuchida N. The p53 tumor-suppressor gene and ras oncogene mutations in oral squamous-cell carcinoma. Int J Cancer. 1992 Dec 2;52(6):867-72. PMID:1459726
- ↑ Groesser L, Herschberger E, Ruetten A, Ruivenkamp C, Lopriore E, Zutt M, Langmann T, Singer S, Klingseisen L, Schneider-Brachert W, Toll A, Real FX, Landthaler M, Hafner C. Postzygotic HRAS and KRAS mutations cause nevus sebaceous and Schimmelpenning syndrome. Nat Genet. 2012 Jun 10;44(7):783-7. doi: 10.1038/ng.2316. PMID:22683711 doi:10.1038/ng.2316
- ↑ Guil S, de La Iglesia N, Fernandez-Larrea J, Cifuentes D, Ferrer JC, Guinovart JJ, Bach-Elias M. Alternative splicing of the human proto-oncogene c-H-ras renders a new Ras family protein that trafficks to cytoplasm and nucleus. Cancer Res. 2003 Sep 1;63(17):5178-87. PMID:14500341
- ↑ Lander HM, Hajjar DP, Hempstead BL, Mirza UA, Chait BT, Campbell S, Quilliam LA. A molecular redox switch on p21(ras). Structural basis for the nitric oxide-p21(ras) interaction. J Biol Chem. 1997 Feb 14;272(7):4323-6. PMID:9020151
- ↑ Williams JG, Pappu K, Campbell SL. Structural and biochemical studies of p21Ras S-nitrosylation and nitric oxide-mediated guanine nucleotide exchange. Proc Natl Acad Sci U S A. 2003 May 27;100(11):6376-81. Epub 2003 May 9. PMID:12740440 doi:10.1073/pnas.1037299100
