Human Glucagon Class B G Protein-Coupled Receptors (GPCRs)
Introduction
Human glucagon class B G protein-coupled receptors (GPCRs), also known as secretin-like receptors, are a subfamily of the more well known class A (rhodopsin-like) glucagon receptor family[1] . Located in the liver, class B glucagon receptors (GCGRs) are activated by the binding of the hormonal peptide glucagon, as seen in Figure 1, which leads to the release of glucose into the bloodstream and plays an essential role in glucose homeostasis. Class B GCGRs are composed of a seven transmembrane domain (7TM) and extracellular domain (ECD) that are of vital importance in glucagon binding. In comparison, class A vs. class B glucagon receptors share less than fifteen percent sequence homology, but both share this 7TM which is a primary area of comparison between the two [1]. The understanding of class A family of GCGRs structure-function mechanism has made great progress over the past few years, but understanding of class B has fallen behind.
Structures of Class A vs. Class B GPCRs
Comparison of the was compared to that of a , and it was found that the orientation and positioning of the alpha helices are conserved through both classes. But, structural alignments of the two revealed multiple gaps in the transmembrane region signifying a variety of structural deviations in transmembrane helices [2]. The N-terminal end of helix one in class B GCGR, located in the 7TM, is longer than any known class A GPCR structure and stretches three supplementary helical turns above the extracellular (EC) membrane boundary. This region is referred to as the stalk and is involved in glucagon binding and helps in defining the orientation of the ECD with respect to the 7TM domain [2]. Also specific to class B GPCRs, a glycine (Gly) residue at position 393 induces a ; this bend is stabilized by the hydrophobic interaction between the . One of the most distinguishable characteristics of the class B 7TM is the helix VIII tilt of 25 degrees compared to that of class A, which has no tilt. This results from a glutamate Glu) 406 in helix VIII that is fully conserved in secretin-like receptors and forms two interhelical salt bridges with conserved residues arginine (Arg) 173 and arginine 346 [2]. Despite these differences, a vital region that is conserved in both class B and class A receptors is the disulphide bond between cysteine in extracellular loop two (ECL2). This bond stabilizes the receptors entire 7TM fold. Lastly, the locations of the extracellular tips for class B glucagon receptors allow for a much wider and deeper binding cavity in the ligand-binding pocket, which is much more immense than any of the class A GCGRs [2]. These wide ranges specifically occur between two sets of alpha helices, as seen in Figure 2 below.
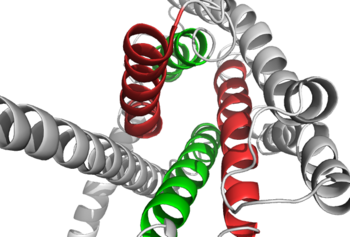
Figure 2: Extracellular tips of the 7TM helices. Helices two and six are shown in green, while helices three and seven are shown in red
How These Structures Lead to Function
Structurally, the N-terminal and the 7TM comprise the signature seven helical structure that is involved in signaling via coupling to heterotrimeric G proteins that activate adenylate cyclase to increase the levels of intracellular cyclic AMP. Additionally, this coupling increases inositol phosphate and intracellular calcium levels [2]. The wider and deeper ligand-binding pocket of class B GPCRs allows for a vast array of receptors to be bound that allow for numerous functions activated by peptide receptors [3]. The conformation and orientation of the 7TM and the ECD regions dictate the functionality of the protein, which has an open and closed conformation of the GCGR. When glucagon binds to GCGR, the open conformation of GCGR is stabilized. There is no clear binding site location of the hormone peptide ligand, but they do know the N-terminus of glucagon binds deep into the . The amino acids at the N-terminus have the ability to form hydrogen bonds and ionic interactions involved, which can be seen in the amino acid sequence of glucagon here in Figure 3. [4].
Many of the residues that are in direct contact with the glucagon molecule are charged or are polar.
There are also many smaller residues on glucagon that support the bulky residues on the GCGR. These residues are located within the of the 7TM [3]. There are specific amino acid interactions that hold the helices of the 7TM in the closed conformation that maximizes affinity. This includes a disulfide bond between mentioned earlier that serves to hold the ECL1 and ECL2 in the proper orientation. Additionally, the salt bridges between Glu 406, Arg 173, and Arg 346, also mentioned earlier, hold the conformation together for higher affinity. Finally, alpha helical structure of the is imperative to the affinity and binding of the glucagon [2].
[[Image:Screen_Shot_2016-03-22_at_5.28.03_PM.png|(|):|425 px|center|thumb|Binding Pocket Residues: side chains of carbon chains are shown in green and side chains containing oxygen are shown in red]
Clinical Relevancy
Of the fifteen human class B GPCRs, eight have been identified as potential drug target[5]. Therapeutic agents have been created from the peptides themselves within this protein, but overall pharmaceutical companies have had difficulty creating agents that act on family B GPCRS. There is an outward appearance and inherent flexibility in the class B GCGR 7TM because of conserved hydrogen bonds that flank a glycine residue, and this structure along with the ECD and its role of interactions on the extracellular side of receptors may provide evidence to how class B receptors adjust its conformational spectra for various receptors. Researchers hope to show how these conformations can be utilized in potential treatments of a wide array disorders.
Potential Inhibitors
Research for class B GCGR inhibitors is primarily looking into allosteric inhibitors having the ability to target specific receptors in order to treat problems like stress disorders, managing hyperglycemia, and also alternative mechanisms for treating migraines [6]. Known inhibitors include monoclonal antibodies which inhibit glucagon receptors through an allosteric mechanism. The monoclonal antibodies bind to two different sites, the ECD opposite of the binding region and then the helical portion of the ECD as well. [7].
Research
Determining the structure of class B GCGRs is a reason for its lack of advanced knowledge in the field, but X-ray crystallography and NMR have been the main processes performed and have had some success with it over the past couple years [8]. X-ray crystallography displayed the crystal structure of ECDs of class B GPCRs in complex with their ligands along with the crystal structure of the 7TM. In addition to this, NMR has allowed the ability to directly understand structures of soluble amino-terminal domains of numerous members of the secretin-like family that bind peptide hormones. Primary sequences analysis have led to the finding of seven segments of eighteen or more relatively hydrophobic residues that are believed to represent transmembrane helices that take part in creating an intramembranous helical bundle [8]. Also, mutagenesis has been used to determine which residues were necessary in maximizing affinity for glucagon. Finally, the orientation and mechanism of the peptide interactions within these structures are studied using peptide structure-activity relationships (SAR), receptor and ligand fragments, chimeric receptors, site-directed mutagenesis, photochemical cross-linking, and molecular modeling [8].


{kind=link}
{kind=link}