Sandbox Reserved 1481
From Proteopedia
| Line 48: | Line 48: | ||
Both MTD have approximatly 25% sequence homology.Despite some common point the methyltransferase domain of METTL14 has extra terminal extensions, with an unusual N-terminal extension which is approximately 50 amino acid long. In the one hand the N-terminal extension create along helix which go through the domain and allow a close contact with the MTD of the catalytic subunit of the complex. This contact is made by several loops and shorter helical segments. In other hand, the C-terminal helix of the MTD of METTL14 is antiparallel to the N-terminal extension helix in order to stabilize its position. This allow the formation of a broad interdomain binding interface,and also others contact point along the terminal extensions, which are stabilizing the positions of both domains neccesary for the function.<Structure load='5YZ9' size='350' frame='true' align='right' caption='The zinc finger domain of METTL3 subunit' scene='Insert optional scene name here' /> | Both MTD have approximatly 25% sequence homology.Despite some common point the methyltransferase domain of METTL14 has extra terminal extensions, with an unusual N-terminal extension which is approximately 50 amino acid long. In the one hand the N-terminal extension create along helix which go through the domain and allow a close contact with the MTD of the catalytic subunit of the complex. This contact is made by several loops and shorter helical segments. In other hand, the C-terminal helix of the MTD of METTL14 is antiparallel to the N-terminal extension helix in order to stabilize its position. This allow the formation of a broad interdomain binding interface,and also others contact point along the terminal extensions, which are stabilizing the positions of both domains neccesary for the function.<Structure load='5YZ9' size='350' frame='true' align='right' caption='The zinc finger domain of METTL3 subunit' scene='Insert optional scene name here' /> | ||
| + | '''Zinc finger binding domain of the METTL3 subunit or the CCCH sequence''' | ||
| - | + | Zinc finger domain of the METTL3 N6-methyladenosine methyltransferase, is a RNA binding domain of the complex. This perticular domain of the two proteins complex allow the protein to make interactions with the RNA molecule to modify through consensus sequences | |
| - | Zinc finger domain of the METTL3 N6-methyladenosine methyltransferase, is a RNA binding domain of the complex. This perticular domain of the two proteins complex allow the protein to make interactions with the RNA molecule to modify | ||
| - | '''The CCCH sequence''' | ||
The CCCH sequences corresponds to a Cys-Cys-Cys-His motif. This specific motif belongs to METTL3 and belongs to the family of zinc binding motifs | The CCCH sequences corresponds to a Cys-Cys-Cys-His motif. This specific motif belongs to METTL3 and belongs to the family of zinc binding motifs | ||
There are two od these CCCH motifs near by the methyltransferase domain of METTL3. As zinc-binding motifs such as CCCHs are known for their abilities to bind nucleic acids such as RNA, their presence are required for RNA methylatransferase activity. In case of a punctual mutation in these sequences a loss of the methyltransferase activity is observed, confirming their importance. | There are two od these CCCH motifs near by the methyltransferase domain of METTL3. As zinc-binding motifs such as CCCHs are known for their abilities to bind nucleic acids such as RNA, their presence are required for RNA methylatransferase activity. In case of a punctual mutation in these sequences a loss of the methyltransferase activity is observed, confirming their importance. | ||
Revision as of 16:54, 30 December 2018
| This Sandbox is Reserved from 06/12/2018, through 30/06/2019 for use in the course "Structural Biology" taught by Bruno Kieffer at the University of Strasbourg, ESBS. This reservation includes Sandbox Reserved 1480 through Sandbox Reserved 1543. |
To get started:
More help: Help:Editing |
Crystal structure of the catalytic domains of Mettl3/Mettl14 complex
|
The complex METTL3/METTL14 is a heterodimer enzymatic complex involved into RNA post-transcriptional modifications by humans. This complex is abble to add a methyl group on adenosin of the RNA, by catalyzing a m6(A) modification.The N(6)-methyladenosine (m(6)A) is a quite common, reversible chemical modification of RNAs molecules, which plays a key role in several different biological fonctions. This post-transcriptional modification can be added by WRITERS, recognized by READERS and also removed byr ERASERS. The METTL3/METTL14 complex plays the role of writer.
This enzymatic complex belongs to the second class of enzymes, which are the transferases. The complex is formed by 574 amino acid residues, divided into two different proteins nammed as Methyltransferase Like number 3 and 14. 
You may include any references to papers as in: the use of JSmol in Proteopedia [1] or to the article describing Jmol [2] to the rescue.
Structural highlights of the METTL3/METTL14 complex
Primary structure
This complex is composed of two differents proteins forming two distinct but linked subunits of the enzymatic complex. Each of these two proteins is coded by one different gene. The mettl3 gene codes for the N6-adenosine-methyltransferase 70 kDa subunit, which is a 225 Amino acid chain, whereas the Mettl14 gene codes for the N6-adenosine-methyltransferase subunit METTL14, which is composed of 349 amino acids.
Each of these two proteins are corresponding to the A [2]or the B chain [3] of the complex and are neccessary to the functionning of the whole complex.
Secondary structure
Both polypeptides METTL3 and METTL14 are able to form some secondary structures thank to interchain and intrachain hydrogen bond formation.
Chain A METTL3'secondary structure, is mainly composed of 20% helical structures (7 helices; 46 residues) and 24% parallel and anti-parallel beta sheets (13 strands; 55 residues)
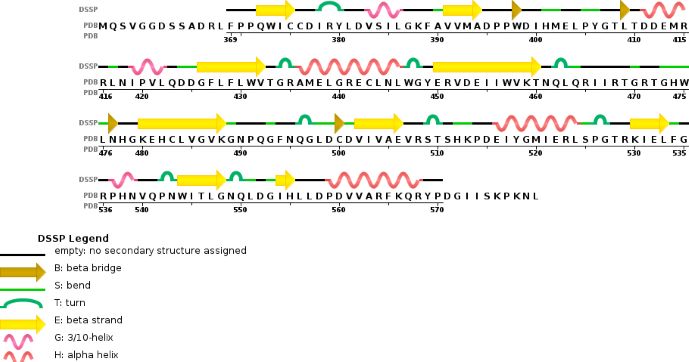

Chain B METTL14 is made of 24% helical structures (14 helices; 87 residues) and 18% parallel and anti-parallel beta sheets (16 strands; 63 residues)
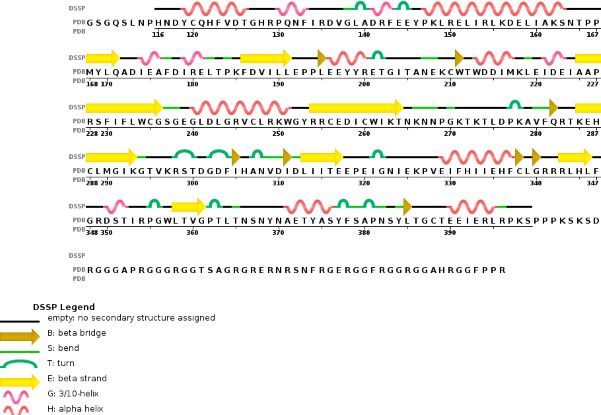

Tertiary structure Thanks to their primary amino acid sequences both A and B chains have some specific conserved domains directly linked to their function and functionning.
Domains and specific site or sequences
MTD : Methyltransferase domain
METTL3 and MTTL14 have both a methyltranferase domain, which is the domain able to catalyze a methyltransferase reaction.But the complex METTL3/METTL14 has a better methyltransferase activity, than one single subunit activity. Moreover, a mutation in the catalytic center of METTL3 inhibits the hole methyltransferase activity of the complex, whereas a mutation in the catalytic center of METTL14 does not. Thus, METTL3 is the catalytic subunit of the complex and METTL14 enhances the methyltransferasese activity by stabilizing the complex structure and binding to messenger RNA by enabling the recognition of its consensus sequence, GGACU, which is the target for m6A modifications.
Both MTD have approximatly 25% sequence homology.Despite some common point the methyltransferase domain of METTL14 has extra terminal extensions, with an unusual N-terminal extension which is approximately 50 amino acid long. In the one hand the N-terminal extension create along helix which go through the domain and allow a close contact with the MTD of the catalytic subunit of the complex. This contact is made by several loops and shorter helical segments. In other hand, the C-terminal helix of the MTD of METTL14 is antiparallel to the N-terminal extension helix in order to stabilize its position. This allow the formation of a broad interdomain binding interface,and also others contact point along the terminal extensions, which are stabilizing the positions of both domains neccesary for the function.
|
Zinc finger binding domain of the METTL3 subunit or the CCCH sequence
Zinc finger domain of the METTL3 N6-methyladenosine methyltransferase, is a RNA binding domain of the complex. This perticular domain of the two proteins complex allow the protein to make interactions with the RNA molecule to modify through consensus sequences
The CCCH sequences corresponds to a Cys-Cys-Cys-His motif. This specific motif belongs to METTL3 and belongs to the family of zinc binding motifs
There are two od these CCCH motifs near by the methyltransferase domain of METTL3. As zinc-binding motifs such as CCCHs are known for their abilities to bind nucleic acids such as RNA, their presence are required for RNA methylatransferase activity. In case of a punctual mutation in these sequences a loss of the methyltransferase activity is observed, confirming their importance.

METTL3 and METTL14 are linked to each other to form the functionnal complex. So as the METTL3 is responsible of the catalytic activity, the METTL14 allow the allosteric recognition of the RNA. The two subunit bindind creates a cavity in the center of the complex where the enzymatic reaction take place, it is the catalytic center. In these catalytic center some critical amino acid residues for the substrate recognition and specific binding are located.
Crystal structure
To study the crystal structure of the METTL3/METTL14 some crystalograghy experiments have been realized under the following experimental conditions to crystalyze the protein complex:
Method : Vapor Diffusion Hanging Drop
pH 8
Temperature : 291.0 K
Buffer: 0.1 M Tris pH 8.0, 20% PEG3350
After crytalization, crystal had been analyzed through X-Ray diffraction, at 100°C, with a single wavelength coming out of a synchrotron. Thanks to the experiment the following data have been collected to describe the cristal structure of the complex :
Unit Cell caracteristics : Length (Å) a = 101.86 b = 101.86 c = 117.66
Angle (°) α = 90 β = 90 γ = 90
Symmetry : Space Group P 41 21 2
The primitive cubic symmetry of the unit cell is driven by two 2-fold axis rotation. Moreover, within this space group which is acentric, chiral, and enantiomorphic with one single lattice translations, eight different representative symmetry operations are needed to get the whole unit cell.

The crystal structure analysis at 1,65 angstrom allow to collect precise and detailled informations about the whole structure like the side chains and also the hydrogen bonds. (Cristal analysis data at other resolution are available on [4].)
Complex METTL3/METTL14 enzymatic working process
The enzymatic complex of two proteins is abble to realize methylation reactions thanks to its sequence and specific which allow it to catalyze some perticular reactions at given sites.
The complex METTL3/METTL14 belongs to the writers group, which means that theses two proteins are taking part to the protein subunit able to realize the post transcriptional modification of the RNA within the humans cells.
The METTL3/METTL14 complex catalyze the methylation reaction. 
As previously said the METTL3 is the catalytic component of the methyltransferase complex. So it is responsible of the m6A modifications thanks to its ability to crosslink with an other essential protein the S-adenosylmethionine (SAM). Moreover, METTL3 can be associated with METTL14, which contains its own methyltransferase domain. Even if the precise molecular interactions between the to subunits are unknown, it is known that the both have to associated with each other, but also that aditionnal factors are neccessary to allow the add of a methyl group on the targeted RNA.
After the formation of the minimal complex with the crystallized heterodimer of methyltransferase domains from METTL3 and METTL14 and two CCCH motifs of Mettl3, and the binding of all the factors the RNA modification reaction can take place.
Function of the modification within the cell
Specific, and well controlled methylation of the nucleic acids is essential for proper gene regulation, whatever the studied organism or modification’s type. However the precise molecular role of m6A is unknown, it is knonw that for most mRNAs, m6A modification appear in long exons, near stop codons, and in 3′ UTRs and that this modification could influence mRNA stability, induce RNA conformational changes, modulate protein-RNA interactions, and even modify microRNA processing One of the most prevalent modifications observed for mRNAs is N6-methyladenosine (m6A). Recent studies have intensely investigated how m6A-modification of RNA contributes to central events in biology. Nonfonctional m6A actors such as writers like METTL3 or METTL14, but also readers, and erasers are oftern linked to problems in self-renewal of stem cells, circadian clock and developmental defects, obesity, synaptic signaling, and cancers.
Disease and problems triggered by unfunctionnal METTL3/METTL14 complex
Relevance
This is a sample scene created with SAT to by Group, and another to make of the protein. You can make your own scenes on SAT starting from scratch or loading and editing one of these sample scenes.
</StructureSection>
References
- ↑ . PMID:216315890657
- ↑ . PMID:216315890657
[1] [2] [3] [4] [5] [6] [7] [8] [9]
POUR URSULA :)
m6A RNA Methylation Controls Neural Development and Is Involved in Human Diseases Kunzhao Du, Longbin Zhang[…]Tao Sun Molecular Neurobiology (2018)
The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells Ly P Vu, Brian F Pickering[…]Michael G Kharas Nature Medicine (2017)
ET CA AUSSI : ca vient de cette publi : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4958592/
In an independent study, the Y406C mutation of Mettl3 was identified in large intestine cancer patients (Colon Adenocarcinoma - TCGA, US (reanalyzed by Cancer Genome Project, Sanger Institute); Study ID: COSU376) (Forbes et al., 2015). Thus, we tested how the Y406C mutation affects methyltransferase activity. When full-length Mettl3 with the Y406C mutation is in complex with wild type Mettl14, we can obtain stable protein complexes but cannot observe any significant level of methylation in vitro (Figures 5D, 5E, S1B and S6D). Loss of the tyrosine ring may prevent Mettl3 from interacting properly with the RNA substrate near the adenine to be methylated and catalyzing the methyl transfer.
In our structures, a highly basic patch is found on MTD14, near the MTD3/MTD14 junction close to the catalytic cavity (Figure 5F). One of these basic residues is the highly conserved arginine 298 of Mettl14, which is frequently mutated to proline in endometrial cancer (Uterine Corpus Endometrioid Carcinoma (TCGA, US) import from ICGC; Study ID: COSU419)(Bell, 2014; Forbes et al., 2015) (Figure S5E). Wild type Mettl3 in complex with full-length Mettl14 with the R298P mutation shows significantly decreased methylation activity (Figures 5D, 5E, S1B and S6D). In addition, these complexes can no longer distinguish the cognate RNA target from the mutant RNA substrate. R298 also lies close to the third “fence” loop discussed above, and may have a complex role to affect both rate and RNA specificity. Thus, Mettl14 seems to play a role in substrate recognition as well as structurally supporting the catalytic cavity in Mettl3.'
