Introduction
Acetylation of lysine residues 5 and 12 found in histones are known to regulate the interconversion of chromatin structures between heterochromatin (inactive) and euchromatin (active) [1]. The role of Histone Acetyltransferase (HAT1) is to transfer an acetyl group from Acetyl-CoA to lysine 5 or lysine 12 on histone IV [2] . This results in an increase in gene activity via transcription activation. The addition of acetyl groups to histones results converts heterochromatin to its more euchromatin form [3]. As acetyl groups are added to lysine, the negatively charged histone begins to lose its negative charge allowing for the more relaxed and active chromatin structures to form. We analyzed the structure and function of the HAT1 gene that’s derived from cerevisiae Saccharomyces cerevisiae[4]. The particular protein model we investigated is called 1BOB and is found on the protein database. The various binding sites and interaction mechanisms involved in the acetylation process were analyzed in addition to the initial Acetyl CoA binding mechanism.
Structure
The structure of HATs consist of a
with at least 3 stranded antiparallel β-sheets and one α-helix spanning the length of the sheet [1]. Specifically, Hat1 has an elongated and curved structure that is comprised of 320 residues. The elongated shape of Hat1 allows for the formation of a concave surface where Acetyl CoA binds to the protein.
Domains
It is composed of a mixture of helices and sheets that form two domains. The domains are connected via an extended loop region that together make up the quaternary structure of the protein. The stretches from residues 2-111 and the
extends from residues 122-320 [3]. Residues 112–121 are what are thought to construct the extended loop that connects the two domains. Additionally, the C-terminal domain contains the active site [3].
Active Site
The concave groove mentioned above is where Acetyl CoA binds and is known to be the active site of the enzyme. The groove contains approximately 1100 Å of accessible surface area [3]. Because Acetyl CoA exhibits a bent shape it is thought that the ligand is able to wrap itself around the protein upon binding. Functional studies state that a conformational change is likely after Acetyl CoA is bound; this conformational change results in the formation of a for the target lysine residue to enter the backside of the active site to be Acetylated [3]. The binding of the target histone does not result in any conformational change. The Lys-12 side chain is able to approach the carbonyl group from the backside of the active site adjacent to the gating region. Near 60% of the Acetyl CoA molecule is found buried in a highly non-polar region of the protein surface called the hydrophobic pocket.
Hydrophobic Pocket
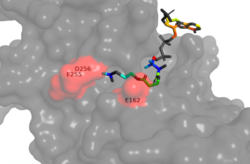
Figure 1. Image of the substrate Acetyl CoA bound in the groove of HAT1 (1BOB.pdb). The groove contains the hydrophobic pocket and is also contains the location of the active site.
The active site consists of the Acetyl CoA ligand bound to the enzyme in a groove on the surface of the protein. The
ligand is held in place by several bonds to protein residues that result in the formation of a . The hydrophobic pocket consists of the interacting side chains from residues in addition to further bonds resulting from residues 217-220 and 255-256 [3] (figure 1). The amide of main-chain
hydrogen bonds to the carbonyl oxygen of the Acetyl group in the binding pocket. The main-chain amide of
also donates a hydrogen bond from its side chain to oxygen PO5 of the pantothenic acid group [3]. The binding within the hydrophobic pocket is further supplemented through the creation of a protein gate that establishes a bridge over the concave surface that serves to keep Acetyl CoA in place while the enzyme interacts with the histone.
Mechanism
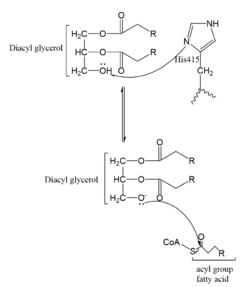
Figure 2. One possible acetylation mechanism of HAT1. Mechanism consists of deprotonation of Lys-12 and then transfer of an acetyl group from Acetyl CoA to Lys-12.
Several mechanisms for the transfer of the acetyl group to the Lys-12 of the histone have been proposed. One possible mechanism consists of a 2-step process. The acetyl group is first transferred to a holding intermediate on the enzyme before subsequently being transferred to Lys-12. There are two cysteine residues, and , that could serve as sites to accept and acetyl group but are not in close enough proximity to the active site
[4]. Another proposed mechanism that has been conserved across several HAT1 studies consists of deprotonating the amino group on the Lys-12 of H4 (figure 2). This mechanism was proposed by Wu et al with respect to the human HAT1 protein. Analysis was done on the
1BOB model (retrieved from the protein database) and similar residues with respect to location and function were identified. Residues , , and cooperate to form a that establishes a basic environment that deprotonates the Lys-12 reside of H4
[4]. Of those residues current research is uncertain which (if any) accept the proton of Lys-12. Deprotonation of the amino group on the Lys-12 makes the residue nucleophilic enough to directly attack the carbonyl carbon to initiate the acetyl transfer
[4]. The transfer mechanism is contingent on the conformational change and the formation of a functional gate that spans the concave groove over the bound Acetyl CoA and holds it in place while the catalytic
deprotonation process takes place.
Protein Gate
The consists of several residues that coordinate a conformational change in the protein to hold the Acetyl CoA ligand in place. , , and cooperate together form the gate over Acetyl CoA [3]. The hydrogen bond between the main chain amide of to the side chain carbonyl oxygen of Asn-258 allows for proper positioning of Asn-258 to bond with other residues and Acetyl CoA itself. The side chain amide of Asn-258 also binds via a water molecule (H2O-415) with the main chain amide of the Glu-162 [3]. These bonds help lock the orientation of Asn-258 so that it can donate a hydrogen bond from its side-chain amide to the carbonyl oxygen PO5 at the end of the pantothenic acid group. Together, the side chains of Ile-161, Glu-162, and Asn-258 form a protein gate over the Acetyl CoA binding cleft. The gate allows for correct binding of Acetyl CoA and once the ligand is bound, subsequent conformational changes of HAT1 happen, allowing for Lys-12 to act as a nucleophile in the precise place.
Homology to Other Proteins
Homology of HAT1 to other proteins was shown by the analysis on similarity of secondary structural elements. The analysis showed that the Hat1 gene is conserved among other organisms, such as the chimpanzee, Rhesus monkey, dog, cow, mouse, rat, chicken, zebrafish, fruit fly, mosquito, C. elegan, K. lactis, E. gossypii, S. pombe, M. oryzae, N. crassa, and A. thaliana [3]. Overall, 269 organisms have orthologs to Hat1. These organisms showed similarity in aligned sequences over much of their length but had variability in the C-terminal domain. A possible reasoning for this is the idea that the C-terminal is not important for the HAT activity of Hat1 [3]. Specifically, these organisms portrayed similar function in multiple histone targets, such as H4, H5, and H12 [3].
References
- ↑ 1.0 1.1 Kleff S, Andrulis ED, Anderson CW, Sternglanz R. Identification of a gene encoding a yeast histone H4 acetyltransferase. J Biol Chem. 1995 Oct 20;270(42):24674-7. PMID:7559580
- ↑ DesJarlais R, Tummino PJ. Role of Histone-Modifying Enzymes and Their Complexes in Regulation of Chromatin Biology. Biochemistry. 2016 Mar 22;55(11):1584-99. doi: 10.1021/acs.biochem.5b01210. Epub , 2016 Jan 26. PMID:26745824 doi:http://dx.doi.org/10.1021/acs.biochem.5b01210
- ↑ 3.00 3.01 3.02 3.03 3.04 3.05 3.06 3.07 3.08 3.09 3.10 3.11 Kleff S, Andrulis ED, Anderson CW, Sternglanz R. Identification of a gene encoding a yeast histone H4 acetyltransferase. J Biol Chem. 1995 Oct 20;270(42):24674-7. PMID:7559580
- ↑ 4.0 4.1 4.2 Wu H, Moshkina N, Min J, Zeng H, Joshua J, Zhou MM, Plotnikov AN. Structural basis for substrate specificity and catalysis of human histone acetyltransferase 1. Proc Natl Acad Sci U S A. 2012 Jun 5;109(23):8925-30. Epub 2012 May 21. PMID:22615379 doi:http://dx.doi.org/10.1073/pnas.1114117109
Student Contributors
Kiran Kaur, Emily Leiderman, Benjamin Nick


