Introduction
Within the central nervous system (CNS), various membrane receptors exist to detect extracellular signaling molecules and communicate this information intracellularly. Found in eukaryotes and known for its seven transmembrane helices, G-protein coupled receptor (GPCR) are one type of membrane bound receptors with conserved intracellular signaling via heterotrimeric G-proteins. The metabotropic glutamate receptor (mGlu), a Class C GPCR, is a receptor utilized in glutamate signaling- which is essential in synaptic plasticity as well as the development and repair of the CNS. These receptors are specifically found in the pre- and postsynaptic neurons of the CNS. Eight different mGlu subtypes exist which are divided into three groups (I, II, III). While each mGlu has a slightly different function and location, the structures of the different mGlu subtypes are very similar (Table 1) [1]. For each group, binding of the neurotransmitter glutamate to the mGlu introduces a conformational change that can activate a G-protein.
| Group
| mGlu Type
| Location
| Function
|
| I
| mGlu1
mGlu5
| Postsynaptic neurons
| Activates calcium channels
Excites adenylyl cyclase
|
| II
| mGlu2
mGlu3
| Presynaptic neurons
| Activates potassium channels
Inhibits calcium channels
Inhibits adenylyl cyclase
|
| III
| mGlu4
mGlu6
mGlu7
mGlu8
| Presynaptic neurons
| Activates potassium channels
Inhibits calcium channels
Inhibits adenylyl cyclase
|
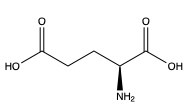
Figure 1. Structure of glutamate.
To activate the mGlu transformation, glutamate acts as the protein's main agonist. Glutamate is an acidic, polar amino acid (Figure 1). This agonist binds to the extracellular portion of the glutamate receptor causing the transmembrane spanning region of the homodimer to change conformationally. This change initiates a signaling cascade within the cell that can ultimately lead to the modification of other proteins and a difference in the synapse’s excitability
[2]. In mGlu, the binding affinity of glutamate is also controlled by the binding of either a positive (PAM) or negative (NAM) allosteric modulator to a binding pocket within the seven TMD
[3].
Structural Highlights
mGlu receptors are dimeric proteins consisting of an . While a heterodimer of different mGlu subtypes can form, only homodimeric receptors can become active[2]. Both the alpha and beta chains are comprised of : the venus fly trap (VFT), cysteine rich domain (CRD), and the transmembrane domain (TMD).
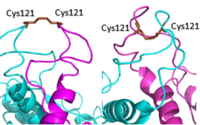
Figure 2. Cysteine 121 positioning in the inactive (left) vs the active (right) mGlu conformation
Domains
: The extracellular location in which the two glutamate agonists bind is known as the VFT. This domain includes a disulfide bond between C121 of the alpha and beta chains. This bond is shifted down in the inactive form and undergoes an upward movement upon glutamate binding which stabilizes the active site (Figure 2).
: The portion of the protomer that connects the VFT with the TMD is known as the CRD. As the connecting segment of the protein, it is critical in transmitting the conformational change caused by the binding of glutamate to the TMD. The change resulting from the binding of glutamate in the VFT brings the cysteine-rich domain together to alter the configuration of the TMD through its interaction with the extracellular loop 2 (ECL2) [2]. This process is mediated by the hydrophobic effect due to the nature of the amino acids at the apex of the CRD [2].
: The TMD consists of seven transmembrane helices that are responsible for G-protein interactions. In the inactive form, the asymmetric conformation of the helices is mediated by the hydrophobicity of helix 3 and 4 [2]. Along with the interaction of the CRD with the ECL2 of the TMD, an allosteric modulator must bind within the transmembrane helices to allow for the conformation of the helices to be altered. This conformation allows for an active dimer interface along helix 6 [3]. The stabilization of this conformation also enables G protein coupling with ICL2, ICL3, TM Helix 3 and the C terminus [3].
Conformational Changes
1. In its resting state, mGlu is in an . In this conformation, the receptor is considered open with an inter-lobe angle of 44 degrees
[2]. The structure has two free glutamate binding sites in the VFT, the CRDs are separated, and the TMD is not interacting with a G protein
[2].
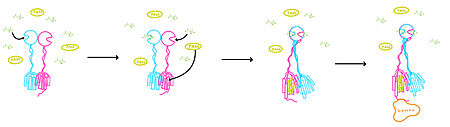
Figure 3. Illustration of mGlu's conformational change process.
2. In the intermediate activation state, also known as the open-closed conformation, one glutamate is bound in one binding pocket of VFT. This single is still considered inactive as the receptor has not changed the conformations in the CRD and thus the TMD. With the same asymmetric transmembrane helices formation, a TM3-TM4 interface is still present and mGlu cannot interact with a G protein[2].
3. A second glutamate then binds to the other of the VFT. Mediated by L639, F643, N735, W773, and F776, a (PAM) also binds within the seven TMD helices of the alpha chain
[2]. This closed conformation with an inter-lobe domain of 25 degrees is considered the active conformation
[2]. The binding of these ligands allows the CRDs to compact and come together. This transformation causes the TMD to form a separate, active asymmetric conformation with a TM6-TM6 interface between the chains
[2].

Figure 4. The interaction between an active mGlu and a G-protein
4. The from the alpha and beta chains allows for intracellular loop 2 (ICL2) and the C-terminus to be properly ordered to interact with a G protein[2]. While hydrogen bonding is present, this coupling is primarily driven by the hydrophobic interactions in the interface with the ɑ5 helix of the G protein [2](Figure 4). This can only occur in the presence of a PAM as the pocket in which the coupling occurs would be completely closed in its absence[2].
5. Depending on the type of mGlu present, different signaling cascades will occur within the cell [3]. These cascades are necessary for cellular function and can lead to various diseases[4].
Clinical Relevance
Glutamate is a major neurotransmitter in the brain and is known for its role in memory, synaptic plasticity, and neuronal development[4]. Thus, mGlus are located throughout the CNS and play key roles in various neurodegenerative diseases such as Alzheimer's Disease (AD), Huntington's Disease (HD), Parkinson’s Disease (PD), and Amyotrophic Lateral Sclerosis (ALS)[4]. Also, due to the key role of glutamate in the brain, mGlus are also being studied for their role in psychiatric disorders like anxiety and depression[4]. In many of these diseases, the overexpression of glutamate can lead to the overstimulation of mGlus and excitotoxicity [5]. However, an exception to this is the under stimulation of mGlus which can result in schizophrenia[6]. Research is ongoing, but studies have proven that agonists, antagonists, PAMs, and NAMs of mGlus are potential treatments for these diseases[4].




