This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
User:Mitchell Long/Sandbox 1
From Proteopedia
| |||||||||||
| |||||||||
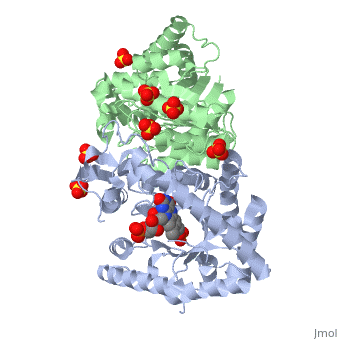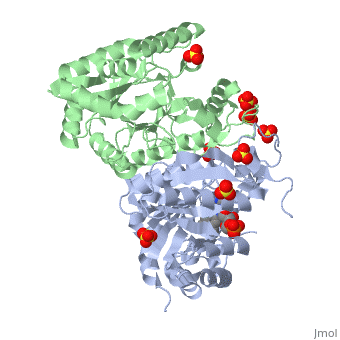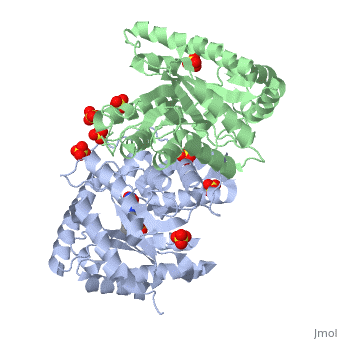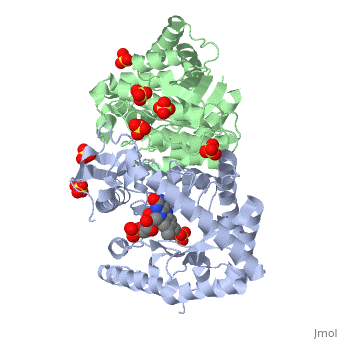
| 3fgc, resolution 2.30Å () | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ligands: | , , | ||||||||
| Gene: | luxA (Vibrio harveyi), luxB (Vibrio harveyi) | ||||||||
| Activity: | Alkanal monooxygenase (FMN-linked), with EC number 1.14.14.3 | ||||||||
| Related: | 1luc, 1brl | ||||||||
| |||||||||
| |||||||||
| Resources: | FirstGlance, OCA, RCSB, PDBsum | ||||||||
| Coordinates: | save as pdb, mmCIF, xml | ||||||||

Applications In Biotechnology
Luciferases are most commonly used as reporter genes by transduction or transfection assays. Reporter genes are inserted into an organism with a gene of interest. This is a powerful method of measuring gene expression because it is non-invasive. Genes for luciferase can paired with an inducible operon. When the gene for luciferase and the gene of interest are incorporated into the host genome, they can "turned on" by induction. Once the desired gene is turned on, gene expression can be determined by the intensity of the light produced by transcription of the gene of interest.
Quorum Sensing
In a process known as quorum sensing, bacteria communicate using secreted signal molecules called autoinducers(AIs). 'V. harveyi' is a mesophilic, gram negative, rod shaped bacteria that can communicate with other bacteria via quorum sensing. Quorum-sensing bacteria alter gene expression in response to the accumulation of AIs, which reflects an increase in cell population density[1]. This process is believed to provide bacteria a means to coordinately control the gene expression of the group, giving them multicellular characteristics. When bacteria reach a "quorum," their population has reached a density high enough to coordinate gene expression[1]. Often, bacteria make and respond to multiple AIs. Vibrio harveyi, a free-living marine bacterium, produces at least three distinct AIs to control bioluminescence, biofilm formation, Type III Secretion (TTS), and protease production. When a bacterial population density is low, the LuxI gene is transcribed constitutively at basal level. The three V. harveyi AIs are HAI-1, an acyl homoserine lactone; AI-2, a furanosyl-borate-diester; and CAI-1, of unknown structure[1]. When the population density reaches an adequate level, the conjugate receptor LuxR begins transcription. LuxR is the regulatory receptor, and when an AI binds the the LuxR receptor, transcription is turned on resulting in the production of more AI and the expression of other genes involved in quorum sensing. When V. harveyi reaches a high enough population density, it's quorum sensing genes are activated and the transcription of the genes that code for the luciferase enzyme. [4] [1] [5]
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 Waters CM, Bassler BL. The Vibrio harveyi quorum-sensing system uses shared regulatory components to discriminate between multiple autoinducers. Genes Dev. 2006 Oct 1;20(19):2754-67. PMID:17015436 doi:10.1101/gad.1466506
- ↑ Campbell ZT, Weichsel A, Montfort WR, Baldwin TO. Crystal Structure of the Bacterial Luciferase:Flavin Complex Provides Insight into the Function of the subunit. Biochemistry. 2009 May 12. PMID:19435287 doi:http://dx.doi.org/10.1021/bi900003t
- ↑ Campbell ZT, Weichsel A, Montfort WR, Baldwin TO. Crystal Structure of the Bacterial Luciferase:Flavin Complex Provides Insight into the Function of the subunit. Biochemistry. 2009 May 12. PMID:19435287 doi:http://dx.doi.org/10.1021/bi900003t
- ↑ Campbell ZT, Weichsel A, Montfort WR, Baldwin TO. Crystal Structure of the Bacterial Luciferase:Flavin Complex Provides Insight into the Function of the subunit. Biochemistry. 2009 May 12. PMID:19435287 doi:http://dx.doi.org/10.1021/bi900003t
- ↑ Fisher AJ, Raushel FM, Baldwin TO, Rayment I. Three-dimensional structure of bacterial luciferase from Vibrio harveyi at 2.4 A resolution. Biochemistry. 1995 May 23;34(20):6581-6. PMID:7756289
