Czr Operon
The Chromosome Determined Zinc Responsible (Czr) operon acts as described above, with Czr A acting as a regulator protein. As such, Czr A is responsible for controlling the transcription of the rest of the operon and by extension, the transport of Zn 2+ out of the cell; the role of Czr A in the Czr operon is described in further detail as part of the explanation of biological function. In addition to being a component of an operon, Czr A is also considered to be a metal sensor protein. While the immediate function of Czr A is gene regulation, this serves the larger purpose of acting to maintain an appropriate concentration of Zn 2+ in the cell.
Biological Function
Czr A is a transcriptional repressor protein responsible for the regulation of the Czr operon[1]. The Czr operon contains genes for the proteins Czr A and Czr B. Czr B is a Zinc transport protein which moves Zn2+ out of a cell while Czr A regulates this process by controlling expression level of Czr B. When relatively low amounts of zinc are present in the cell Czr A will bind to DNA, preventing the progression of RNA polymerase and thus inhibiting expression of Czr B. Decreased expression of Czr B results in the ability of the cell to retain Zn2+ more readily. Because Czr A and Czr B are transcribed as part of the same operon, an inhibitor of Czr A must be readily available to allow full transcription of Czr B when necessary. Czr A is noncompetitively inhibited by the binding of two Zn2+ ions, which is ideal in that this allows expression of Czr B, a Zn2+ transporter to be dependent on the relative amount of Zn2+ in the cell. Czr A displays two different conformations; the first binds DNA and has relatively low affinity for Zn2+ (PDB code: 2kjb). In this conformation the . Binding of zinc drives a conformational change (PDB code: 2kjc) in which the , lowering the affinity for DNA.
Zinc Binding
Zinc acts as an inhibitor to Czr A[1], thus preventing transcriptional repression of Czr B and allowing Zn2+ transport out of the cell. This allows for zinc transport to essentially be self regulated. That is, when zinc concentration in the cell is relatively high, zinc ions bind to Czr A, causing a conformational change which releases the bound DNA. DNA without Czr A bound is free to be transcribed and Czr B is again expressed, allowing for Zn2+ transport out of the cell. At low Zn2+ concentrations, CzrA represses RNA Polymerase activity, and Zn2+ ions are maintained inside the cell.
Structural Overview
CzrA functions as a dimer. The dimerize at the czr operon, repressing gene transcription. Each monomeric unit contains seen in purple and displayed in yellow. While the function of the beta sheets are not yet known, key helices regulate the binding of DNA and Zn+2. The are the location of DNA binding and the contain the Zn+2 binding site. As Zn+2 ions binds the alpha 4 helices are , repressing their DNA binding ability (Figure 2). Two seperate PDB codes exist for CzrA: CzrA with DNA bound (2KJB) and CzrA with zinc+2 bound (2KJC). Unfortunately, zinc ions are not visible in the 2KJC NMR structure that was obtained for CzrA.
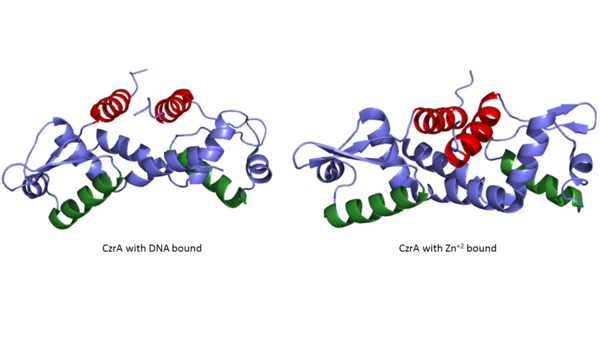
Figure 2:Comparison of CzrA with Zn
+2 bound and CzrA with DNA bound with the alpha five helices shown in red and the alpha four helices shown in green
DNA Binding
Ser 54, Ser 57, and His 58 have been found to be the main . are likely to interact with the 5'-TGAA sequence found in the half-site of the DNA, where the alpha 4 helices . These residues are found in the N terminal of the alpha 4 helix (figure 3). Additionally, Val 42 and Gln 53 are involved in the . This conclusion was experimentally determined by mutagenesis of the Gln and Val with Ala residues and measuring the mutant DNA binding capacity. In a previously published article [1], the DNA bound state of Czr A was tested by using the known critical residues for DNA interactions. Gln 53, Val 42 (red), Ser 54, Ser 57, and His 58 (orange) were individually mutated to Ala, and kinetic experiments were performed. Compared to wild type Czr A, mutating Gln53 and V42 residues resulted in an 11-fold and 160-fold decrease in Ka, respectively. Mutations to the main DNA interaction sites Ser 54, Ser 57, and His 58 resulted in binding similar to the , suggesting that these residues are essential to binding DNA. While the conformational change that occurs from the Zinc to DNA bound state of Czr A is small,the alpha 4 helices (shown in green in Figure 2) are slightly shifted. The loss of DNA binding in the mutagenesis experiements in combination with the lack of any other major physical changes between these two states further supports that the alpha 4 helices are the location of DNA binding in Czr A. Experimental data can be found in table 1 from this same article.
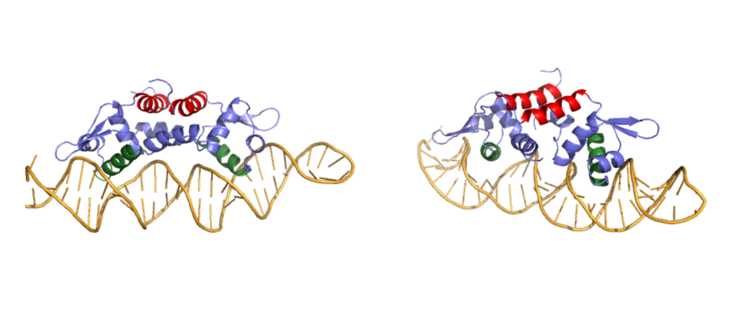
Figure 3:Side by side rotation showing the main DNA groove in CzrA
Zinc Binding
Many zinc-dependent proteins are transcriptional regulators[2]. Czr A fits into this category as an allosteric inhibitor of the czr operon. Two Zn +2 ions may bind to the dimer[1], at the location of the helix from each monomer. As zinc binds, the alpha 5 helices to inhibit the DNA binding residues (Figure 2). Furthermore, CzrA must be in its dimer form for zinc to bind. The is formed by two residues from each monomer, so Zn+2 cannot bind to the monomer. The is formed by Asp 84 and His 86 from one monomer, and His 97 and His 100 from the other monomer. Zinc ions were not present in the solution NMR crystal structure, so a representation of a zinc ion in the binding pocket can be seen in figure 4. Histidine residues are a repetitive and commonly found residue in zinc-binding proteins [3].
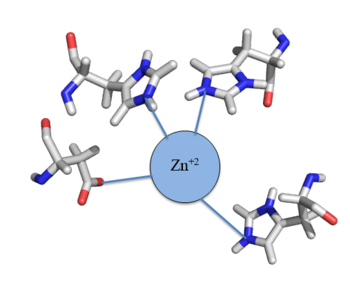
Figure 4:Zn
+2 tetrahedral binding complex
Zinc+2 binding is driven by a large entropic gain [4]. Water molecules around the metal ion and CzrA protein are displaced, and gain greater freedom. This gain in entropy allows Zn+2 to bind to CzrA with reasonable affinity and speed in vivo. The zinc+2 ion forms a tetrahedral complex with the four residues (Figure 3). This allows other metal ions to act as allosteric inhibitors to CzrA. Any metal that may form a tetrahedral complex will have some affinity for CzrA, assuming it is not too large to fit into the pocket. However, the metal binding pocket of CzrA has been optimized to bind Zn+2 with the highest affinity. As CzrA is a transcriptional repressor, binding of Zn+2 to the dimer will activate the czr operon. Zn+2 is preferred as CzrB opens a Zn+2 channel, allowing the excess zinc ions to export the cell.




