Introduction
Vitamin K epoxide reductase (VKOR) is the enzyme responsible for regenerating vitamin K from vitamin K epoxide to support blood coagulation.
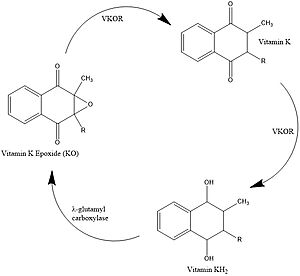
Figure 1. Vitamin K Cycle
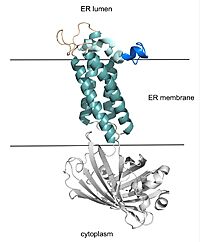
Figure 2. VKOR with Barrel Domain
Vitamin K Cycle
Vitamin K is essential for blood clotting in the body. The fully reduced form, KH2, allows the gamma carboxylation of blood clotting cofactors and is turned into the epoxide form in the process. Vitamin K epoxide reductase, abbreviated VKOR, turns the epoxide back to the fully reduced form so the reduced form can be used again. This transformation happens in two steps including converting the epoxide to the partially oxidized Vitamin K quinone then converting the quinone to the fully reduced hydroquinone (KH2). [1]
Structural Overview
VKOR consists of four embedded in the endoplasmic reticulum membrane. The files on the RCSB Protein Data Bank include a barrel domain that is not pertinent to the function of VKOR. The images presented here have been edited to remove the barrel domain and renumbered to correspond with the article by Liu. [2]. The original image with the barrel domain in context is shown in Figure 2. Helices one and two are connected by the region which contains two of the active cysteines, C43 and C51. VKOR also has a covering the active site, made up of an , , and .
Catalytic Cycle
Overview
There are 4 catalytic cysteines that are important to VKOR, 43, 51, 132, and 135. To explain how this works, it is easiest to start with the second state. In the second state, an oxidized or partially oxidized Vitamin K has entered the active site. The stabilizing 51-132 disulfide bond is shown. Then in the third state, 43 has attacked the disulfide bond and made its own bond with 51. You can see 132 has an oxygen. That is because the researchers made a mutation from S to O to force the reaction to stop at that step so the structure could be deduced. In the natural VKOR, that would be a sulfur. The next state, the open state, results from 132 forming a bridge with 135. This allows release of the reduced or partially reduced Vitamin K. All of this disulfide rearranging was working to reduce the Vitamin K, particularly in the 135 position. If we go back to State 2, when Vitamin K first binds, you can see that 135 is not tied up in a disulfide bond. It is available to help the Vitamin K bond. So, it makes sense that once 135 gets forced to bond, the now reduced Vitamin K is released. State 5 is interesting because the disulfide bonds are similar to the open state, but warfarin is actually bound. This represents the binding of warfarin to the fully oxidized VKOR at the end of its cycle. Going back to State 1, the researchers used a mutation at 43 to mimic VKOR’s partially oxidized state. Warfarin can also bind to this state and notice that the disulfide bonds are the same as State 2. Also it is worth pointing out how the disulfide bonds contribute to conformational changes and are affected by conformational changes, which affects their proximity to each other and the active site.
Catalytic Cysteines
VKOR uses four catalytic cysteines (43, 51, 132, and 135) to facilitate reduction and cause conformational changes via disulfide bridge formation. When an oxidized or partially oxidized Vitamin K enters the active site, VKOR has a stabilizing C51-C132 disulfide bond, known as the closed state. C135 is not in a disulfide bridge and is important in helping Vitamin K bind. C43 attacks the C51-C132 bond, forming a new C43-C51 bond that characterizes the open state. The free amino acid at the 132 position is shown as a serine in this depiction because it was mutated it to stop the reaction at this intermediate during Liu's experimental procedure. [2]. In the wild type VKOR, the 132 position would be a cysteine. C132 forms a bridge with C135 which allows release of the reduced or partially reduced Vitamin K. These rearrangements facilitate the use of cysteines as reducing agents for Vitamin K. Cysteine rearrangements also contribute to and are affected by overall conformational changes which affects their proximity to each other and the active site.
Warfarin binding also depends on the catalytic cysteines. Warfarin is able to bind to the fully oxidized form of VKOR where the disulfide bridge pairings are C132-C135 and C43-C51. Warfarin can also bind to the partially oxidized form of VKOR where the disulfide bridge pairings are the same as the closed state, with C51-C132 being the only bridge. Again, C43 is shown as a serine because a mutation was used to force VKOR to adopt that conformation during the experimental procedure by Liu. [2].
Catalytic Amino Acids
VKOR uses two catalytic amino acids, tyrosine 139 and asparagine 80, to stabilize in all forms and , such as warfarin, in the binding pocket. Tyr139 and Asn80 hydrogen bond to carbonyl groups on both structures and stabilizes them within the binding pocket.
Hydrophobic Interactions
Medical Relevance
Warfarin
Warfarin is a structural mimic of Vitamin K that is used clinically as an anticoagulant.
Superwarfarins


