B-cell lymphoma protein
From Proteopedia
Contents |
This is a placeholder
This is a placeholder text to help you get started in placing a Jmol applet on your page. At any time, click "Show Preview" at the bottom of this page to see how it goes.
Replace the PDB id (use lowercase!) after the STRUCTURE_ and after PDB= to load and display another structure.
| |||||||||
| 3cin, resolution 1.70Å () | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ligands: | , , | ||||||||
| Gene: | TM1419, TM_1419 (Thermotoga maritima MSB8) | ||||||||
| Activity: | Inositol-3-phosphate synthase, with EC number 5.5.1.4 | ||||||||
| |||||||||
| |||||||||
| Resources: | FirstGlance, OCA, RCSB, PDBsum, TOPSAN | ||||||||
| Coordinates: | save as pdb, mmCIF, xml | ||||||||
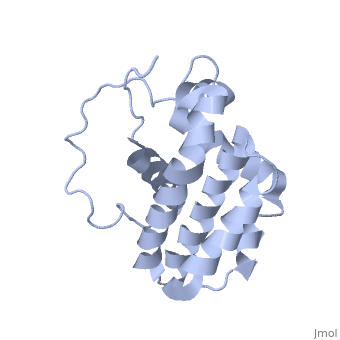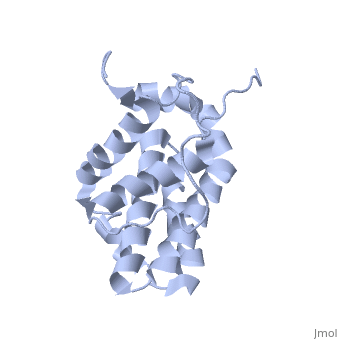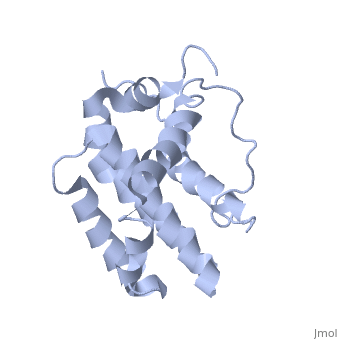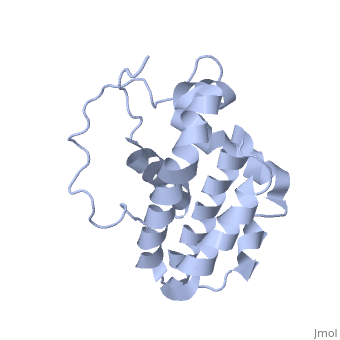

Function (I)
There are a number of theories concerning how the Bcl-2 gene family exert their pro- or anti-apoptotic effect. An important one states that this is achieved by activation or inactivation of an inner mitochondrial permeability transition pore, which is involved in the regulation of matrix Ca2+, pH, and voltage. It is also thought that some Bcl-2 family proteins can induce (pro-apoptotic members) or inhibit (anti-apoptotic members) the release of cytochrome c into the cytosol which, once there, activates caspase-9 and caspase-3, leading to apoptosis. Although Zamzami et al. suggest that the release of cytochrome c is indirectly mediated by the PT pore on the inner mitochondrial membrane,[1] strong evidence suggest an earlier implication of the MAC pore on the outer membrane.[2][3] Bcl-2 family[4]
Another theory suggests that Rho proteins play a role in Bcl-2, Mcl-1 and Bid activation. Rho inhibition reduces the expression of anti-apoptotic Bcl-2 and Mcl-1 proteins and increases protein levels of pro-apoptotic Bid but had no effect on Bax or FLIP levels. Rho inhibition induces caspase-9 and caspase-3-dependent apoptosis of cultured human endothelial cells.[5]
The members of the Bcl-2 family share one or more of the four characteristic domains of homology entitled the Bcl-2 homology (BH) domains (named BH1, BH2, BH3 and BH4) (see the figure on the left). The BH domains are known to be crucial for function, as deletion of these domains via molecular cloning affects survival/apoptosis rates. The anti-apoptotic Bcl-2 proteins, such as Bcl-2 and Bcl-xL, conserve all four BH domains. The BH domains also serve to subdivide the pro-apoptotic Bcl-2 proteins into those with several BH domains (e.g. Bax and Bak) or those proteins that have only the BH3 domain (e.g. Bid, Bim and Bad). The Bcl-2 family has a general structure that consists of a hydrophobic helix surrounded by amphipathic helices. Many members of the family have transmembrane domains. The site of action for the Bcl-2 family is mostly on the outer mitochondrial membrane. Within the mitochondria are apoptogenic factors (cytochrome c, Smac/DIABLO, Omi) that if released activate the executioners of apoptosis, the caspases.[6] Depending on their function, once activated, Bcl-2 proteins either promote the release of these factors, or keep them sequestered in the mitochondria. Whereas the activated pro-apoptotic Bak and/or Bax would form MAC and mediate the release of cytochrome c, the anti-apoptotic Bcl-2 would block it, possibly through inhibition of Bax and/or Bak.[7]
Role in disease
The Bcl-2 gene has been implicated in a number of cancers, including melanoma, breast, prostate, and lung carcinomas, as well as schizophrenia and autoimmunity. It is also thought to be involved in resistance to conventional cancer treatment. This supports a role for decreased apoptosis in the pathogenesis of cancer.
Cancer is one of the world's leading causes of death and occurs when the homeostatic balance between cell growth and death is disturbed. Research in cancer biology has discovered that a variety of aberrations in gene expression of anti-apoptotic, pro-apoptotic and BH3-only proteins can contribute to the many forms of the disease. An interesting example can be seen in lymphomas. The over-expression of the anti-apoptotic Bcl-2 protein in lymphocytes alone did not act in an oncogenic manner. But simultaneous overexpression of Bcl-2 and the protooncogene myc may produce aggressive B-cell malignancies including lymphoma.[8] In follicular lymphoma, a chromosomal translocation commonly occurs between the fourteenth and the eighteenth chromosomes—t(14;18)—which places the Bcl-2 gene next to the immunoglobulin heavy chain locus. This fusion gene is deregulated, leading to the transcription of excessively high levels of bcl-2.[9] This decreases the propensity of these cells for undergoing apoptosis.
Apoptosis also plays a very active role in regulating the immune system. When it is functional, it can cause immune unresponsiveness to self-antigens via both central and peripheral tolerance. "In the case of defective apoptosis, it may contribute to etiological aspects of autoimmune diseases.[10] The autoimmune disease, type 1 diabetes can be caused by defective apoptosis, which leads to aberrant T cell AICD and defective peripheral tolerance. Due to the fact that dendritic cells (DCs) are of the most important antigen presenting cells of the immune system, their activity must be tightly regulated by such mechanisms as apoptosis. "Researchers have found that mice containing DCs that are Bim -/-, thus unable to induce effective apoptosis, obtain autoimmune diseases more so than those that have normal DCs.[10] Other studies have shown that the lifespan of DCs may be controlled by factors such as a timer dependent on anti-apoptotic Bcl-2.[10] These investigations illuminate the importance of regulating antigen presentation as mis-regulation can lead to autoimmunity.
Apoptosis plays a very important role in regulating a variety of diseases that have enormous social impacts. For example, schizophrenia is a neurodegenerative disease that may result from an abnormal ratio of pro- and anti-apoptotic factors.[11] There is some evidence that this defective apoptosis may result from abnormal expression of Bcl-2 and increased expression of caspase-3.[11]
Further research into the family of Bcl-2 proteins will provide a more complete picture on how these proteins interact with each other to promote and inhibit apoptosis. An understanding of the mechanisms involved will help discover potential treatments such as inhibitors to target over-expressed proteins that may lead to new therapies in cancer, autoimmune conditions, and neurological diseases.
Targeted Therapies
An antisense oligonucleotide drug Genasense (G3139) has been developed to target Bcl-2. An antisense DNA or RNA strand is non-coding and complementary to the coding strand (which is the template for producing respectively RNA or protein). An antisense drug is a short sequence of RNA which hybridises with and inactivates mRNA, preventing the protein from being formed.
It was shown that the proliferation of human lymphoma cells (with t(14;18) translocation) could be inhibited by antisense RNA targeted at the start codon region of Bcl-2 mRNA. In vitro studies led to the identification of Genasense, which is complementary to the first 6 codons of Bcl-2 mRNA.[12]
These have shown successful results in Phase I/II trials for lymphoma, and a large Phase III trial is currently underway[13]
Genasense did not receive FDA approval after disappointing results in a melanoma trial.
Abbott has recently described a novel inhibitor of Bcl-2, Bcl-xL and Bcl-w, known as ABT-737.[14] ABT-737 is one among many so-called BH3 mimetic small molecule inhibitors (SMI) targeting Bcl-2 and Bcl-2-related proteins such as Bcl-xL and Bcl-w but not A1 and Mcl-1, which may prove valuable in the therapy of lymphoma and other blood cancers.[15]
Proteopedia Page Contributors and Editors (what is this?)
Michal Harel, Colin Ridenour, David Canner, Alexander Berchansky, Jaime Prilusky
