Hepatocyte growth factor receptor
From Proteopedia
Contents |
Hepatocyte Growth Factor Receptor
| |||||||||||
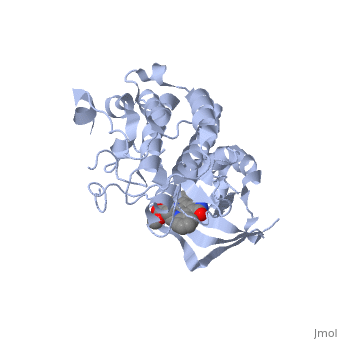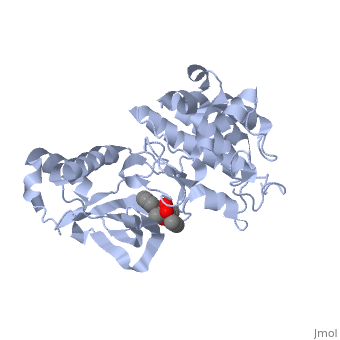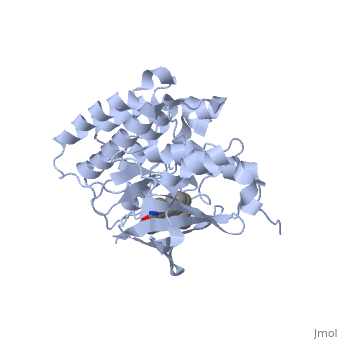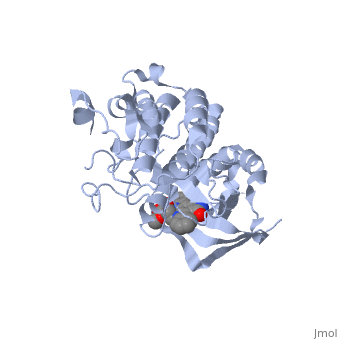
Mutated Receptor in Complex with K-252a
|
Mutation
This particular structure of the hepatocyte growth factor tyrosine kinase domain is one harboring a human cancer mutation. The two are replaced by a phenylalanine and aspartate, respectively. This mutation normally causes the receptor to be consitutively active, and is found in metastatic HNSC carcinoma. Although there is no longer phosphorylation at these sites, it is believed that the aspartate negative charge resembles the negative phosphate that would normally cause activation, and therefore keeps the protein in its active form. [10] There is a third mutation at Tyr-1194 which is substituted for a . This is shown to point into the formed by Lys-1198 and Leu-1195 from αE. [11] This structure is conserved in the wild type protein, suggesting that the mutation at residue 1149 is not changing the structure at this position.
K-252a
is a staurosporine analog. Staurosporine is an inhibitor of many Ser/Thr Kinases, and has been shown to also inhibit c-Met activation by inhibiting its autophosphorylation. The structures of K-252a and staurosporine are very similar, with the main difference being that K-252a has a furanose instead of a pyranose Moiety. The binding of K-252a causes the c-Met to adopt an inhibitory conformation of the A-loop, specifically with residues . This segment blocks the place where the substrate tyrosine side chain would bind, if the protein were in an active conformation. Residues also enhance this inhibitory conformation, as they constain C into a conformation that does not allow the catalytic placement of Glu-1127, and by inhibiting nucleotide binding. The K-252a itself binds in the adenosine pocket, therefore inhibiting the binding of ATP. The binding of K-252a is very favorable (enthalpy change of -17.9 kcal/mol). this is probably due to polar interactions as well as a change in conformation upon binding. [12]
There is a concerteted conformational change in the complex upon K-252a binding. One of these changes involves the A-loop, specifically residues 1228-1230. In the Apo-Met structure, the side chain of Met-1229 would pass through the ring of the inhibitor, and so, in order to make room for K-252a, the segment must move, with residues 1229 and 1230 moving by 3-4 Å. In order to make room for the side chain of Tyr-1230, arg-1208 moves by 8 Å toward Asp-1204. Are-1208, which in the uninhibited complex would stack with tyr-1230, now stacks with phe-1234 [13]
K-252a binds in the adenosine pocket. It has four hydrogen bonds to the enzyme, with of these mimicking hydrogen bonds of an adenine base. There is a hydrogen bond between the lactam nitrogen and the carbonyl oxygen of Pro-1158, and another between the lactam carbonyl oxygen and the hydrogen of the amide of Met-1160. There are two more hydrogen bonds between the 3' hydroxyl and carbonyl oxygen and the of the A loop. [14]
There are also many hydrophobic interactions between the interface of the enzyme and K-252a. The residues involved in this are Ile-1084, Gly-1085, Phe-1089, Val-1092, Ala-1108, Lys-1110, and Leu-1140 (); Leu-1157, Pro-1158, Tyr-1159, and Met-1160 (); and Met-1211, Ala-1226, Asp-1228, Met-1229, and Tyr-1230 (). [15]
Met-1229, Met-1211 and Met-1160 all make up the for the indolocarbazole plane as they are all within van der waals distance of it. [16]
C-Terminal Docking Site
There are two binding motifs of the docking site, 1356YVNV and 1349YVHV, each containing a tyrosine that gets phosphorylated. These tyrosines correspond to residues . In this strcuture, as mentioned above, these tyrosine residues have been substituted for phenylalanine and so do not become phosphorylated. Because of this, residues form an extended conformation, while residues 1353-1356 and 1356-1359 form .
Biological Significance
Many human cancers can be traced back to mutations in the c-met kinase domain, and these mutations often lead to over activation of this kinase. Many of the mutations affect the A loop conformation of the wild type receptor, causing it to become constitutively active. This is done by either stabilizing the active form of the enzyme or destabilizing the inactive form. This structure shows that the A loop is flexible and will adapt in order to bind to an indolocarbazole. This gives insight on designs for specific c-Met inhibitors, as this site can be targeted for binding of the inhibitor.
References
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ http://en.wikipedia.org/wiki/C-Met
- ↑ Maina F, Casagranda F, Audero E, Simeone A, Comoglio PM, Klein R, Ponzetto C. Uncoupling of Grb2 from the Met receptor in vivo reveals complex roles in muscle development. Cell. 1996 Nov 1;87(3):531-42. PMID:8898205
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
- ↑ Schiering N, Knapp S, Marconi M, Flocco MM, Cui J, Perego R, Rusconi L, Cristiani C. Crystal structure of the tyrosine kinase domain of the hepatocyte growth factor receptor c-Met and its complex with the microbial alkaloid K-252a. Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12654-9. Epub 2003 Oct 14. PMID:14559966 doi:10.1073/pnas.1734128100
