Introduction
The insulin receptor is a transmembrane receptor [1] that resides in the plasma membrane and is activated by the binding of insulin. The insulin receptor belongs to the large class of receptor tyrosine kinase (RTKs). RTKs are found at the cell surface and have a high affinity for a particular ligand. RTKs are made up of three distinct parts: an extracellular domain with ligand binding sites, a transmembrane region, and an intracellular domain with the tyrosine kinases that initiate intracellular signaling cascades [1]. Downstream signaling from the insulin receptor initiates a variety of cellular pathways including glucose homeostasis, regulation of lipid, protein, and carbohydrate metabolism, gene expression, and even modulation of brain neurotransmitter levels [2]. Amongst RTKs, the insulin receptor is unusual as its undergoes a large conformation change upon insulin binding. Through recent cryo-EM structures of the insulin receptor bound in various conformations, a complete three-dimensional understanding of this conformational changes in finally coming into focus. This page focuses specifically on the insulin receptor's role in glucose homeostasis.
Structural Overview
The insulin receptor is a dimer of heterodimers made of two and two [2]. Within the extracellular ectodomain, there are four potential that can interact with insulin ligands on the extracellular side of the membrane. The insulin receptor has only been imaged in pieces, which has led to proposed structures of the entirety of the receptor based on the function of downstream tyrosine autophosphorylation.https//www.rcsb.org/structure/1ir3 PDB 1IR3
Alpha Subunits
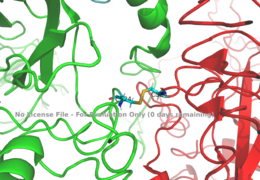
Figure 1: Disulfide bridge (yellow) made of two cysteine residues (blue) that provides a linkage and stability to the two alpha subunits.
PDB 6SOFThe alpha subunits make up the extracellular domain (ectodomain) of the insulin receptor and are the sites of insulin binding. The alpha subunit is comprised of two Leucine rich domains (L1 & L2), a Cysteine rich domain (CR), and a C-terminal alpha helix. The alpha subunits are held together by a disulfide bond between at the CYS524 position on each alpha subunit. The actual site of insulin binding occurs at the of one of the sites discussed next and is stabilized by the L1 and L2 domains. Two types of insulin binding sites are present in the alpha subunits, and . Due to structural differences and accessibility in these binding sites, the first two sites, 1 and 1', have much higher affinity than that of sites 2 and 2'. The sites are in pairs because of the heterodimeric nature of the receptor. Each time an insulin ligand binds to sites 1 and 1', it comes in contact with the L1 domain of one protomer and the alpha-CT chain and FnIII-1 loop of another protomer, which is also known as "cross linking". Insulin can also bind at sites 2 and 2', but the location on the back of the beta sheet of the FnIII-1 domain and lower surface area decreases their binding occupancy. [3] Cryo-EM has imaged insulin bound structures that displayed a T-shape conformation in the alpha subunits.[3]
Beta Subunits
The beta subunit spans from the extracellular domain across the transmembrane region and into the intracellular portion of the insulin receptor. The beta subunit is composed of part of fibronectin domain III-2 and all of Fibronectin domain III-3. The beta subunit's FnIII-3 domain has links through the transmembrane region into the intracellular part of the membrane.Cryo-EM results have displayed clear representations of FnIII-2 and FnIII-3 domains, but lack in their ability to model the receptor structure throughout the transmembrane region and intracellular region. Due to FnIII-3 domain’s connection to these regions, it is proposed that the T-shape conformation extends all the way to the tyrosine kinase domain region.
Function
The insulin receptor's structure is critical to it's function. The receptor begins the signaling pathway that will eventually move glucose transporters to the cell surface which will allow glucose to passively defuse into the cell.
Activation by Insulin
is a
peptide hormone produced and secreted from the
islets of Langerhans of the pancreas in response to high blood glucose levels. Insulin is commonly considered the anabolic hormone of the body, and is the an important
ligand in glucose homeostasis. The structure of insulin is a simple monomer composed of two peptide chains linked by an intermolecular disulfide bridge. The glucose receptor is inactive in the absence of insulin. When there is a surplus of glucose circulating in the blood stream, the production of insulin is upregulated and will bind to many insulin receptors. Upon activation, the receptor undergoes a structural
conformation change from the inactive state to the active state. The activation and conformation change lead to downstream signaling by the phosphorylation of the
Insulin Receptor Substrate (IRS), resulting in glucose intake. The transport of extracellular glucose into the cell allows it to be converted to
glycogen for storage and later usage.
Conformation Change
Structures of the inactive inverted V conformation only contains a single because the entire inactive alpha subunit dimer has been unable to be photographed because the transition state has yet to be determined in full. In the V-shape, the FnIII-3 domains are separated by about 120Å which keeps the tyrosine kinase domains separated. In the V-shape, autophosphorylation and downstream signaling cannot be initiated. Upon the binding of insulin to binding sites 1, 1', and either 2 or 2', the conformation change will begin and bring the FnIII-3 domains within 40Å of each other to induce the T-state conformation. [4] [3] The T shape conformation is well observed in the alpha subunit. It is horizontally composed of L1, CR (including the alpha-CT chain), and L2 domains and vertically composed of the FnIII-1, 2, and 3 domains. This structural transition will facilitate the autophosphorylation of the tyrosine kinase domain.
Binding interactions
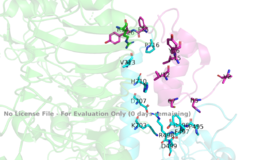
Figure 3: Subunit interactions between the insulin receptor CT-alpha helix (light blue) and insulin (magenta) in one of the binding sites.
PDB 6SOFA tripartite interaction occurs between the alpha-CT chain and the FnIII-1 domain region during the conformational change. This interaction involves the following residues: and the . This duo then interacts with the leucine rich region, L1, that exists on the opposing protomer of the dimer. The tripartite interaction between the alpha-CT chain and FnIII-1 domain on one dimer and the L1 region on the other dimer is important because it allows for a strong and stable interaction between two dimers of the insulin receptor that maintains the T-shape activation state for the rest of the downstream signaling to occur.
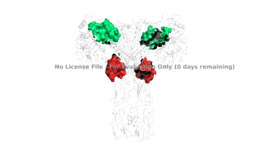
Figure 4: The presence of all four potential binding sides on the active insulin receptor: sites 1 and 1' (green) and sites 2, and 2'(red).
PDB 6SOFFor insulin binding, it is generally more common for only one or two insulin molecules to bind to the receptor due to the occurrence of negative affinity at the binding site. The location of the second two binding sites are on the back side of the Beta subunits and lack favorable surface area. The binding of insulin to the binding sites 1 and 1', as well as one insulin to either binding site 2 or 2', is required for the activation of the insulin receptor and the change in conformation to the active T state. [5].
Although interactions at all four binding sites are highly hydrophobic, the ligand binding interactions at sites 1 and 1' are different than at sites 2 and 2'. Sites 1 and 1' have two disulfide bond linkages, along with HIS B5 from insulin interacting with residues from the FnIII-1 domain. At sites 2 and 2' the FnIII-1 region has interacting with numerous residues on the surface of the insulin ligand.
Relevance
In a healthy individual, insulin is secreted in response to high blood glucose levels is bound by insulin receptors throughout the body which initiates a robust physiological response. Without properly functioning insulin receptors that can respond to increases in insulin, and therefore glucose, medical intervention is necessary for survival.
Disease
Diabetes mellitus is one of the most common diseases involving the insulin receptor in regards to glucose uptake and homeostasis. Type 1 diabetes is classified as "insulin dependent" and involves an inability for the body to produce insulin, resulting from damage or insufficiency in the Islets of Langerhans in the pancreas. Type 2 diabetes is classified as "insulin independent" and is the result of the body producing insufficient amounts of insulin, or not responding to the insulin. There are many molecular mechanisms that cause type 2 diabetes including lipotoxicity, hyperglycemia, and hyperinsulinemia which all cause hyper-activation of various protein phosphatases which reduces phosphorylation of the insulin receptor [6]. Type 1 diabetes is often treated with insulin injections, and type 2 diabetes is treated with diet and lifestyle changes [7] [8].
At the Cellular Level
The conformation change from the inactive to active state upon insulin binding has a time constant of six minutes. Autophosphorylation of the beta subunits reaches a maximal level in about one minute, and lasts for approximately six to ten minutes. One insulin receptor substrate has a half-life of 3.5 minutes where it is able to be phosphorylated by the tyrosine kinases of the beta subunit and then act as a central hub for further downstream signaling pathways. Once insulin binds to the alpha subunit, the receptor remains active for approximately ten minutes before the insulin is degraded and the receptor returns to its inactive conformation. This time frame puts a perspective on how long it takes for the human body to store excess glucose in their blood stream from a recent meal as glycogen for later use as fuel. [2]




