From Proteopedia
proteopedia linkproteopedia link
The FpgNei Protein Superfamily
Background on DNA Repair
The genome of any living organism is continuously affected by exogenous and endogenous agents, such as ultraviolet light, ionizing radiation, different chemicals and the cell's own metabolites (such as reactive oxygen). Therefore, different systems have evolved to repair these damages, with some of these systems shared throughout all lifeforms. Therefore, the proper functioning of DNA repair is critical for survival. There are six pathways of DNA repair (reviewed in Friedberg et al), which include base-excision repair. The latter's distinguishing feature is that it removes lesions as single bases, as opposed to dNMPs or short oligonucleotides like other systems. [1]
Base excision repair's signature enzyme are the DNA glycosylases. These enzymes work by recognizing a damaged base, and then hydrolizing the N-glycosidic bond of the damaged deoxynucleoside and thus removing a single damaged base from DNA. The subsequent steps of the pathway (strand incision, gap-filling and ligation) are done by other enzymes. [2][3]
Background on Fpg Nei
Overall Function and Structure
| (For FPG, the structure used was 1r2y and for Nei 1k3w) Members of this family have . When enzyme binds to DNA, the damaged base
. This superfamily is also characterized by containing a ; as well as a
. Both the H2TH motif and the Zinc finger, as well as other residues
. [4]. Catalysis is believed to be mediated by . For information on the mechanism please consult [5][6][7][8][9].
Similarly In EcoNei (structure 1k3w, , and (Thymine Glycol in the case of Nei). [10]. Some of these amino acids are stabilized by a (although a zincless finger motif is present in some of these subfamilies [11]). Analogously to Fpg, ), Note that the last two elements discussed, the zinc vs. zincless finger, and the two kinds of intercalation loops, are examples of coevolving functional clusters, groups of amino acids that perform a function, and that might be unnecessary or compensated for within the other subfamilies. We developed a novel method for identifying these clusters and have applied it to bring insight into the structure, function and evolution FpgNei family.
|
|
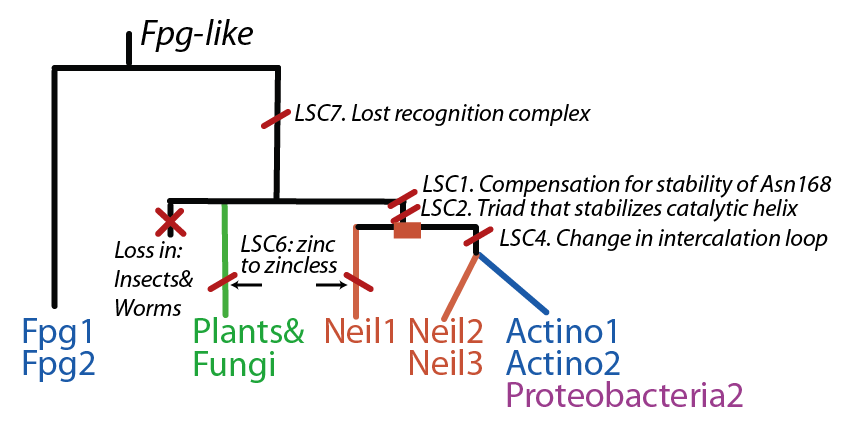
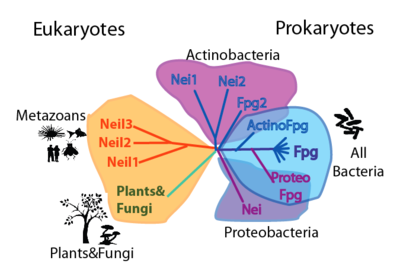 Cartoon phylogenetic tree of the FpgNei protein family. Note that this phylogeny can be appreciated in two levels: by the distribution and number of FpgNei subfamilies in different organisms; and by the kinds of damages that can be repaired |
| Clade | Function | References
|
| Fpg | 8oxoG, Fapy-A, Fapy-G,Me-Fapy-G,Sp,Gh | [12][13]
|
| EcoNei | Oxidized pyrimidines | [14]
|
| Actinomycetes Nei1 | DHU | [15]
|
| Actinomycetes Nei2 | ? |
|
| Fpg2 | ? |
|
| Plant&Fungi | Sp,Gh | [16]
|
| Neil1 | Sp,Gh,on double&single strand DNA | [17]
|
| Neil2 | Sp,Gh,on double&single stranded DNA | [18]
|
| Neil3 | not clear | [19]
|
Several authors have suggested mechanisms for these enzymes, please see references for more information [20][21][22][23].
Homologous structures have been solved, including Fpg protein from Lactococcus Lactis (1pjj)[24], Bacillus Stereothermophilus (1r2y)[25], Thermos Thermophilus (1ee8)[26] and Escherichia Coli(1k82)[27] and Nei from Escherichia Coli (1k3w)[28]. The overall structure is similar, and some of the damages include 8-oxoguanine and fapyG (1xc8)[29].
|
Latent Structural Characters (LSCs)
We define a latent structural character (LSC) as neighboring amino acids which have changed in rate or constraint for a given clade or set of clades. For example, the cysteines in the zinc finger are all conserved in the fpg1, fpg2, actinobacterial and eukaryotic clades, but have a higher rate in plants and neil1, as the latter have a zinc-less finger (see here for more information). We developed a novel method to find these groups of amino acids, discovering previously unknown groups of amino acids which are functionally important in some of these subfamilies.
The table below has some of our main findings, there are two variants for each LSC in case that for one LSC there is a compensating cluster in the other subfamilies. If you click on one of them you will see the result in the structure, and you can go to the explanation and the sequence distribution below.
Suggestion: Once you have an animation you would like to study, we recommend clicking the "pop-up" button to create a new screen so that you can read the text alongside. Click here for Jmol tutorial.
LSC1: Stability of perfectly conserved Asn168
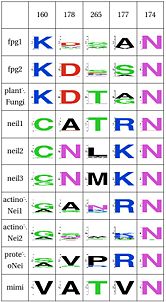 WebLogo for the first functional unit, note the equivalent roles between lysine and arginine, which hydrogen-bond with the crucial Asn174 |
|
|
|
LSC2: Stability of catalytic helix
( and )  The LER triad interact and might stabilize the catalytic helix (which includes Pro2 and Glu3 and Glu6), it is not clear how this is achieved in the other subfamilies |
|
|
|
LSC3: Neil1-specific: Stability of Lys60
 Notice the covariation in Neil1 and the rest with positions 134 and 137 and 170 |
|
|
|
LSC4: Intercalation Loop
and )
 The residue in positions 77 and 78 suggest a possible intercalation loop |
|
|
|
LSC5: DNA Binding Tyrosine
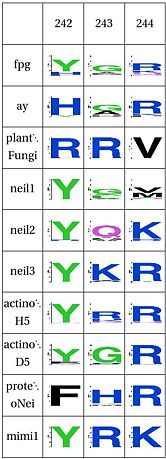 Tyrosine that binds to DNA, plants have a different structure alltogether
This tyrosine hydrogen bonds with Glu179 (BstFpg coordinates). Glu179 is perfectly conserved throughout all clades, and its mutation results in loss of activity [47]. We find that this Tyrosine is replaced by an arginine in plants, however, this arginine could be fulfilling the same role as the tyrosine and might not account for any difference in function. This is consistent with Glu179 and Tyr243 being part of the same network and for Tyr243 playing an important role in the function of the protein. However, this amino acid hasn't been studied. On the other hand, Arg244 is very hightly conserved in everything but Neil1 and plants, consistent with the idea that it might contribute to Zinc Finger stability. The presence of Arg244 correlates with the presence of Pro248, which is the amino acid right before Cys249, which holds the Zinc.
|
|
LSC6: Zinc/zincless finger
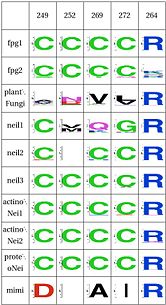 The Zinc Finger helps as a support of the key R264 residue The Zinc finger (of the β/β-antiparallel CCCC type) serves to hold the absolutely conserved Arg264 residue [48], which binds to the phosphate of the damaged base and is crucial for function (mutation of this site results in failed cleavage of the damaged base [49]). Site directed mutagenesis experiments have been performed on all four cysteines, leading to loss of activity [50][51], emphasizing the importance of this LSC. In Neil1, there is no Zinc but there is an equivalent structure: a "zincless finger" [52]. Both the plants and mimivirus have a zincless finger as well [53][54], although it is not clear if these are all homologous.
|
|
|
|
| Functional Cluster | Variant 1 | Variant 2 | Fpg1 | Fpg2 | Plant | Neil1 | Neil2 | Neil3 | Proteo | Actino1 | Actino2 | MimiVirus
|
| Support for perfectly conserved Asn174
|
|
| 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0
|
| Stability of catalytic helix
|
|
| 1 | 1 | 1 | - | - | - | - | - | - | -
|
| Neil1-specific: Stability of Lys60
|
|
| Y | Y | Y | N | Y | Y | Y | Y | Y | Y
|
| Intercalation loop
|
|
| Y | Y | Y | N | N | N | N | N | N | Y
|
| PlantFungi-specific: R254 DNA binding
|
| different in plants
| Y | Y | N | Y | Y | Y | N | Y | Y | Y
|
| Zinc Finger
|
|
| Y | Y | N | N | Y | Y | Y | Y | Y | N
|
| Recognition complex
|
| none
| Y | N | N | N | N | N | N | N | N | N
|
|
|
LSC7: Recognition Complex
This complex is key in recognizing damaged guanine, in the literature it is referred to as the alpha10-b9 loop. For details and hypothesis on the structural basis of 8oxoG and fapyG recognition please refer to [55][56]. The area corresponding to this complex is much shorter in some of the other subfamilies, and there is no obvious homology. In the EcoNei and Neil1 structures this loop is absent as well [57][58]. We showed that the distribution of the rates of substitution within this loop is non-random, providing further support for the importance of the loop in the specificity in Fpg. This non-randomness is also present in 3 of the other subfamilies perhaps suggesting a possible important role of this area in those subfamilies.
|
|
Zinc/zincless finger
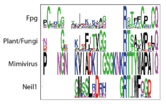
Tyrosine that binds to DNA, the plant has a different structure alltogether
Evolution
The FpgNei evolution has not been easy to resolve [59], especially in the deeper branches. Assuming that functional clusters evolve more slowly than individual residues, we can use this as phylogenetic characters to 1) draw the most parsimonious evolution of the superfamily as dictated by these functional clusters 2) examine how these clusters have evolved and how this might have influenced the evolution of FpgNei.
Site-directed mutants
The following we intend to be comprehensive list of site-directed mutants from the literature. The description is just a very brief, and thus inaccurate summary. Please see the relevant reference for more information on assays used and precise results.
| Name | Organism/Structure | Effect | Reference
|
| | E. coli Nei (1k3w) | Inactive but can still make Schiff Base. | [60][61][62][63]
|
| | E. coli Nei (1k3w) | inactive | [64][65]
|
| | E. coli Nei (1k3w) | Decrease glycosylase, lyase ok. | [66]
|
| | E. coli Nei (1k3w) | Decrease glycosylase, lyase ok. | [67]
|
| | E. coli Fpg (1k82) | No activity. | [68]
|
| | E. coli Nei (1k3w) | No change. | [69]
|
| | E. coli Fpg (1k82) | 3-fold decreased activity. | [70][71]
|
| | E. coli Fpg (1k82) | higher Kd (reduced binding) | [72]
|
| | E. coli Fpg (1k82) | higher Kd (reduced binding) | [73]
|
| | E. coli Fpg (1k82) | similar activity | [74]
|
| | E. coli Fpg (1k82) | reduced glygosylase, less efficient in Schiff-base complex. No cleavage. | [75][76][77][78][79]
|
| | E. coli Fpg (1k82) | similar activity but reduced turnover on 8oxoG:C | [80]
|
| | B. stereo Fpg (1r2y) | Switches preferences for syn and anti in 8oxoG | [81]
|
| | E. coli Nei (1k3w) | Decreased or no activity | [82]
|
| | E. coli Fpg (1k82) | reduced rate of excision | [83]
|
| | E. coli Nei (1k3w) | Active but different kinetics. | [84]
|
| | E. coli Fpg (1k82) | No change. | [85]
|
| | E. coli Fpg (1k82) | Abolished binding | [86]
|
| | E. coli Fpg (1k82) | Decreases processivity | [87]
|
| | E. coli Nei (1k3w) | same | [88]
|
| | E. coli Fpg (1k82) | 20-fold decreased activity. | [89][90]
|
| | E. coli Fpg (1k82) | reduced glygosylase, less efficient in Schiff-base complex. No cleavage. | [91][92]
|
| | E. coli Nei (1k3w) | same | [93]
|
| | E. coli Fpg (1k82) | same | [94]
|
| | E. coli Nei (1k3w) | reduced activity, loss of opposite base discrimination | [95]
|
| | E. coli Nei (1k3w) | ok lyase, reduced glycosylase | [96]
|
| | E. coli Fpg (1k82) | No activity. | [97]
|
| | E. coli Nei (1k3w) | Decreased activity on 5S,6R Tg, slightly less active on DHU | [98]
|
| | B. stereo Fpg (1r2y) | Selectively reduced excision ability | [99]
|
| | E. coli Nei (1k3w) | inactive, although can cleave AP sites | [100]
|
| | H. Sapiens Neil1 (1tdh) | Low activity | [101]
|
| | E. coli Fpg (1k82) | No binding nor cleavage | [102]
|
| | E. coli Fpg (1k82) | No binding nor cleavage. No zinc, no altered secondary structure | [103]
|
| | E. coli Nei (1k3w) | Loss of activity | [104]
|
| | E. coli Fpg (1k82) | No lyase nor glycosylase | [105]
|
|
|
References
- ↑ Zharkov DO. Base excision DNA repair. Cell Mol Life Sci. 2008 May;65(10):1544-65. PMID:18259689 doi:10.1007/s00018-008-7543-2
- ↑ Zharkov DO. Base excision DNA repair. Cell Mol Life Sci. 2008 May;65(10):1544-65. PMID:18259689 doi:10.1007/s00018-008-7543-2
- ↑ Robertson AB, Klungland A, Rognes T, Leiros I. DNA repair in mammalian cells: Base excision repair: the long and short of it. Cell Mol Life Sci. 2009 Mar;66(6):981-93. PMID:19153658 doi:10.1007/s00018-009-8736-z
- ↑ Gilboa R, Zharkov DO, Golan G, Fernandes AS, Gerchman SE, Matz E, Kycia JH, Grollman AP, Shoham G. Structure of formamidopyrimidine-DNA glycosylase covalently complexed to DNA. J Biol Chem. 2002 May 31;277(22):19811-6. Epub 2002 Mar 23. PMID:11912217 doi:http://dx.doi.org/10.1074/jbc.M202058200
- ↑ Pereira de Jesus K, Serre L, Zelwer C, Castaing B. Structural insights into abasic site for Fpg specific binding and catalysis: comparative high-resolution crystallographic studies of Fpg bound to various models of abasic site analogues-containing DNA. Nucleic Acids Res. 2005 Oct 20;33(18):5936-44. Print 2005. PMID:16243784 doi:http://dx.doi.org/33/18/5936
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Coste F, Ober M, Carell T, Boiteux S, Zelwer C, Castaing B. Structural basis for the recognition of the FapydG lesion (2,6-diamino-4-hydroxy-5-formamidopyrimidine) by formamidopyrimidine-DNA glycosylase. J Biol Chem. 2004 Oct 15;279(42):44074-83. Epub 2004 Jul 10. PMID:15249553 doi:10.1074/jbc.M405928200
- ↑ Gilboa R, Zharkov DO, Golan G, Fernandes AS, Gerchman SE, Matz E, Kycia JH, Grollman AP, Shoham G. Structure of formamidopyrimidine-DNA glycosylase covalently complexed to DNA. J Biol Chem. 2002 May 31;277(22):19811-6. Epub 2002 Mar 23. PMID:11912217 doi:http://dx.doi.org/10.1074/jbc.M202058200
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Leipold MD, Muller JG, Burrows CJ, David SS. Removal of hydantoin products of 8-oxoguanine oxidation by the Escherichia coli DNA repair enzyme, FPG. Biochemistry. 2000 Dec 5;39(48):14984-92. PMID:11101315
- ↑ David SS, Williams SD. Chemistry of Glycosylases and Endonucleases Involved in Base-Excision Repair. Chem Rev. 1998 May 7;98(3):1221-1262. PMID:11848931
- ↑ Jiang D, Hatahet Z, Melamede RJ, Kow YW, Wallace SS. Characterization of Escherichia coli endonuclease VIII. J Biol Chem. 1997 Dec 19;272(51):32230-9. PMID:9405426
- ↑ Sidorenko VS, Rot MA, Filipenko ML, Nevinsky GA, Zharkov DO. Novel DNA glycosylases from Mycobacterium tuberculosis. Biochemistry (Mosc). 2008 Apr;73(4):442-50. PMID:18457574
- ↑ Kathe SD, Barrantes-Reynolds R, Jaruga P, Newton MR, Burrows CJ, Bandaru V, Dizdaroglu M, Bond JP, Wallace SS. Plant and fungal Fpg homologs are formamidopyrimidine DNA glycosylases but not 8-oxoguanine DNA glycosylases. DNA Repair (Amst). 2009 May 1;8(5):643-53. Epub 2009 Feb 12. PMID:19217358 doi:10.1016/j.dnarep.2008.12.013
- ↑ Hailer MK, Slade PG, Martin BD, Rosenquist TA, Sugden KD. Recognition of the oxidized lesions spiroiminodihydantoin and guanidinohydantoin in DNA by the mammalian base excision repair glycosylases NEIL1 and NEIL2. DNA Repair (Amst). 2005 Jan 2;4(1):41-50. PMID:15533836 doi:10.1016/j.dnarep.2004.07.006
- ↑ Hailer MK, Slade PG, Martin BD, Rosenquist TA, Sugden KD. Recognition of the oxidized lesions spiroiminodihydantoin and guanidinohydantoin in DNA by the mammalian base excision repair glycosylases NEIL1 and NEIL2. DNA Repair (Amst). 2005 Jan 2;4(1):41-50. PMID:15533836 doi:10.1016/j.dnarep.2004.07.006
- ↑ Takao M, Oohata Y, Kitadokoro K, Kobayashi K, Iwai S, Yasui A, Yonei S, Zhang QM. Human Nei-like protein NEIL3 has AP lyase activity specific for single-stranded DNA and confers oxidative stress resistance in Escherichia coli mutant. Genes Cells. 2009 Feb;14(2):261-70. Epub 2008 Jan 15. PMID:19170771 doi:10.1111/j.1365-2443.2008.01271.x
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Pereira de Jesus K, Serre L, Zelwer C, Castaing B. Structural insights into abasic site for Fpg specific binding and catalysis: comparative high-resolution crystallographic studies of Fpg bound to various models of abasic site analogues-containing DNA. Nucleic Acids Res. 2005 Oct 20;33(18):5936-44. Print 2005. PMID:16243784 doi:http://dx.doi.org/33/18/5936
- ↑ Gilboa R, Zharkov DO, Golan G, Fernandes AS, Gerchman SE, Matz E, Kycia JH, Grollman AP, Shoham G. Structure of formamidopyrimidine-DNA glycosylase covalently complexed to DNA. J Biol Chem. 2002 May 31;277(22):19811-6. Epub 2002 Mar 23. PMID:11912217 doi:http://dx.doi.org/10.1074/jbc.M202058200
- ↑ Pereira de Jesus K, Serre L, Zelwer C, Castaing B. Structural insights into abasic site for Fpg specific binding and catalysis: comparative high-resolution crystallographic studies of Fpg bound to various models of abasic site analogues-containing DNA. Nucleic Acids Res. 2005 Oct 20;33(18):5936-44. Print 2005. PMID:16243784 doi:http://dx.doi.org/33/18/5936
- ↑ Fromme JC, Verdine GL. DNA lesion recognition by the bacterial repair enzyme MutM. J Biol Chem. 2003 Dec 19;278(51):51543-8. Epub 2003 Oct 1. PMID:14525999 doi:10.1074/jbc.M307768200
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Gilboa R, Zharkov DO, Golan G, Fernandes AS, Gerchman SE, Matz E, Kycia JH, Grollman AP, Shoham G. Structure of formamidopyrimidine-DNA glycosylase covalently complexed to DNA. J Biol Chem. 2002 May 31;277(22):19811-6. Epub 2002 Mar 23. PMID:11912217 doi:http://dx.doi.org/10.1074/jbc.M202058200
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Coste F, Ober M, Carell T, Boiteux S, Zelwer C, Castaing B. Structural basis for the recognition of the FapydG lesion (2,6-diamino-4-hydroxy-5-formamidopyrimidine) by formamidopyrimidine-DNA glycosylase. J Biol Chem. 2004 Oct 15;279(42):44074-83. Epub 2004 Jul 10. PMID:15249553 doi:10.1074/jbc.M405928200
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Rabow L, Venkataraman R, Kow YW. Mechanism of action of Escherichia coli formamidopyrimidine N-glycosylase: role of K155 in substrate binding and product release. Prog Nucleic Acid Res Mol Biol. 2001;68:223-34. PMID:11554299
- ↑ Kropachev KY, Zharkov DO, Grollman AP. Catalytic mechanism of Escherichia coli endonuclease VIII: roles of the intercalation loop and the zinc finger. Biochemistry. 2006 Oct 3;45(39):12039-49. PMID:17002303 doi:10.1021/bi060663e
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Pereira de Jesus K, Serre L, Zelwer C, Castaing B. Structural insights into abasic site for Fpg specific binding and catalysis: comparative high-resolution crystallographic studies of Fpg bound to various models of abasic site analogues-containing DNA. Nucleic Acids Res. 2005 Oct 20;33(18):5936-44. Print 2005. PMID:16243784 doi:http://dx.doi.org/33/18/5936
- ↑ Sugahara M, Mikawa T, Kumasaka T, Yamamoto M, Kato R, Fukuyama K, Inoue Y, Kuramitsu S. Crystal structure of a repair enzyme of oxidatively damaged DNA, MutM (Fpg), from an extreme thermophile, Thermus thermophilus HB8. EMBO J. 2000 Aug 1;19(15):3857-69. PMID:10921868 doi:http://dx.doi.org/10.1093/emboj/19.15.3857
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Perlow-Poehnelt RA, Zharkov DO, Grollman AP, Broyde S. Substrate discrimination by formamidopyrimidine-DNA glycosylase: distinguishing interactions within the active site. Biochemistry. 2004 Dec 28;43(51):16092-105. PMID:15610004 doi:10.1021/bi048747f
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Sidorkina OM, Laval J. Role of lysine-57 in the catalytic activities of Escherichia coli formamidopyrimidine-DNA glycosylase (Fpg protein). Nucleic Acids Res. 1998 Dec 1;26(23):5351-7. PMID:9826758
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Perlow-Poehnelt RA, Zharkov DO, Grollman AP, Broyde S. Substrate discrimination by formamidopyrimidine-DNA glycosylase: distinguishing interactions within the active site. Biochemistry. 2004 Dec 28;43(51):16092-105. PMID:15610004 doi:10.1021/bi048747f
- ↑ Kropachev KY, Zharkov DO, Grollman AP. Catalytic mechanism of Escherichia coli endonuclease VIII: roles of the intercalation loop and the zinc finger. Biochemistry. 2006 Oct 3;45(39):12039-49. PMID:17002303 doi:10.1021/bi060663e
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ O'Connor TR, Graves RJ, de Murcia G, Castaing B, Laval J. Fpg protein of Escherichia coli is a zinc finger protein whose cysteine residues have a structural and/or functional role. J Biol Chem. 1993 Apr 25;268(12):9063-70. PMID:8473347
- ↑ Tchou J, Michaels ML, Miller JH, Grollman AP. Function of the zinc finger in Escherichia coli Fpg protein. J Biol Chem. 1993 Dec 15;268(35):26738-44. PMID:8253809
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Kathe SD, Barrantes-Reynolds R, Jaruga P, Newton MR, Burrows CJ, Bandaru V, Dizdaroglu M, Bond JP, Wallace SS. Plant and fungal Fpg homologs are formamidopyrimidine DNA glycosylases but not 8-oxoguanine DNA glycosylases. DNA Repair (Amst). 2009 May 1;8(5):643-53. Epub 2009 Feb 12. PMID:19217358 doi:10.1016/j.dnarep.2008.12.013
- ↑ Bandaru V, Zhao X, Newton MR, Burrows CJ, Wallace SS. Human endonuclease VIII-like (NEIL) proteins in the giant DNA Mimivirus. DNA Repair (Amst). 2007 Nov;6(11):1629-41. Epub 2007 Jul 12. PMID:17627905 doi:10.1016/j.dnarep.2007.05.011
- ↑ Fromme JC, Verdine GL. DNA lesion recognition by the bacterial repair enzyme MutM. J Biol Chem. 2003 Dec 19;278(51):51543-8. Epub 2003 Oct 1. PMID:14525999 doi:10.1074/jbc.M307768200
- ↑ Coste F, Ober M, Carell T, Boiteux S, Zelwer C, Castaing B. Structural basis for the recognition of the FapydG lesion (2,6-diamino-4-hydroxy-5-formamidopyrimidine) by formamidopyrimidine-DNA glycosylase. J Biol Chem. 2004 Oct 15;279(42):44074-83. Epub 2004 Jul 10. PMID:15249553 doi:10.1074/jbc.M405928200
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Doublie S, Bandaru V, Bond JP, Wallace SS. The crystal structure of human endonuclease VIII-like 1 (NEIL1) reveals a zincless finger motif required for glycosylase activity. Proc Natl Acad Sci U S A. 2004 Jul 13;101(28):10284-9. Epub 2004 Jul 1. PMID:15232006 doi:10.1073/pnas.0402051101
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Kuznetsov SV, Sidorkina OM, Jurado J, Bazin M, Tauc P, Brochon JC, Laval J, Santus R. Effect of single mutations on the structural dynamics of a DNA repair enzyme, the Escherichia coli formamidopyrimidine-DNA glycosylase--a fluorescence study using tryptophan residues as reporter groups. Eur J Biochem. 1998 Apr 15;253(2):413-20. PMID:9654091
- ↑ Saparbaev M, Sidorkina OM, Jurado J, Privezentzev CV, Greenberg MM, Laval J. Repair of oxidized purines and damaged pyrimidines by E. coli Fpg protein: different roles of proline 2 and lysine 57 residues. Environ Mol Mutagen. 2002;39(1):10-7. PMID:11813291
- ↑ Sidorkina OM, Laval J. Role of the N-terminal proline residue in the catalytic activities of the Escherichia coli Fpg protein. J Biol Chem. 2000 Apr 7;275(14):9924-9. PMID:10744666
- ↑ Golan G, Zharkov DO, Feinberg H, Fernandes AS, Zaika EI, Kycia JH, Grollman AP, Shoham G. Structure of the uncomplexed DNA repair enzyme endonuclease VIII indicates significant interdomain flexibility. Nucleic Acids Res. 2005 Sep 6;33(15):5006-16. Print 2005. PMID:16145054 doi:http://dx.doi.org/33/15/5006
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Perlow-Poehnelt RA, Zharkov DO, Grollman AP, Broyde S. Substrate discrimination by formamidopyrimidine-DNA glycosylase: distinguishing interactions within the active site. Biochemistry. 2004 Dec 28;43(51):16092-105. PMID:15610004 doi:10.1021/bi048747f
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Sidorkina OM, Laval J. Role of lysine-57 in the catalytic activities of Escherichia coli formamidopyrimidine-DNA glycosylase (Fpg protein). Nucleic Acids Res. 1998 Dec 1;26(23):5351-7. PMID:9826758
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Kuznetsov SV, Sidorkina OM, Jurado J, Bazin M, Tauc P, Brochon JC, Laval J, Santus R. Effect of single mutations on the structural dynamics of a DNA repair enzyme, the Escherichia coli formamidopyrimidine-DNA glycosylase--a fluorescence study using tryptophan residues as reporter groups. Eur J Biochem. 1998 Apr 15;253(2):413-20. PMID:9654091
- ↑ Rogacheva M, Ishchenko A, Saparbaev M, Kuznetsova S, Ogryzko V. High resolution characterization of formamidopyrimidine-DNA glycosylase interaction with its substrate by chemical cross-linking and mass spectrometry using substrate analogs. J Biol Chem. 2006 Oct 27;281(43):32353-65. Epub 2006 Aug 22. PMID:16928690 doi:10.1074/jbc.M606217200
- ↑ Saparbaev M, Sidorkina OM, Jurado J, Privezentzev CV, Greenberg MM, Laval J. Repair of oxidized purines and damaged pyrimidines by E. coli Fpg protein: different roles of proline 2 and lysine 57 residues. Environ Mol Mutagen. 2002;39(1):10-7. PMID:11813291
- ↑ Harbut MB, Meador M, Dodson ML, Lloyd RS. Modulation of the turnover of formamidopyrimidine DNA glycosylase. Biochemistry. 2006 Jun 13;45(23):7341-6. PMID:16752923 doi:10.1021/bi052383p
- ↑ Song K, Hornak V, de Los Santos C, Grollman AP, Simmerling C. Computational analysis of the mode of binding of 8-oxoguanine to formamidopyrimidine-DNA glycosylase. Biochemistry. 2006 Sep 12;45(36):10886-94. PMID:16953574 doi:10.1021/bi060380m
- ↑ Kropachev KY, Zharkov DO, Grollman AP. Catalytic mechanism of Escherichia coli endonuclease VIII: roles of the intercalation loop and the zinc finger. Biochemistry. 2006 Oct 3;45(39):12039-49. PMID:17002303 doi:10.1021/bi060663e
- ↑ Zaika EI, Perlow RA, Matz E, Broyde S, Gilboa R, Grollman AP, Zharkov DO. Substrate discrimination by formamidopyrimidine-DNA glycosylase: a mutational analysis. J Biol Chem. 2004 Feb 6;279(6):4849-61. Epub 2003 Nov 7. PMID:14607836 doi:10.1074/jbc.M310262200
- ↑ Dizdaroglu M, Burgess SM, Jaruga P, Hazra TK, Rodriguez H, Lloyd RS. Substrate specificity and excision kinetics of Escherichia coli endonuclease VIII (Nei) for modified bases in DNA damaged by free radicals. Biochemistry. 2001 Oct 9;40(40):12150-6. PMID:11580290
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Zaika EI, Perlow RA, Matz E, Broyde S, Gilboa R, Grollman AP, Zharkov DO. Substrate discrimination by formamidopyrimidine-DNA glycosylase: a mutational analysis. J Biol Chem. 2004 Feb 6;279(6):4849-61. Epub 2003 Nov 7. PMID:14607836 doi:10.1074/jbc.M310262200
- ↑ Sidorenko VS, Zharkov DO. Correlated cleavage of damaged DNA by bacterial and human 8-oxoguanine-DNA glycosylases. Biochemistry. 2008 Aug 26;47(34):8970-6. Epub 2008 Aug 2. PMID:18672903 doi:10.1021/bi800569e
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Perlow-Poehnelt RA, Zharkov DO, Grollman AP, Broyde S. Substrate discrimination by formamidopyrimidine-DNA glycosylase: distinguishing interactions within the active site. Biochemistry. 2004 Dec 28;43(51):16092-105. PMID:15610004 doi:10.1021/bi048747f
- ↑ Rabow LE, Kow YW. Mechanism of action of base release by Escherichia coli Fpg protein: role of lysine 155 in catalysis. Biochemistry. 1997 Apr 22;36(16):5084-96. PMID:9125531 doi:10.1021/bi963005a
- ↑ Rabow L, Venkataraman R, Kow YW. Mechanism of action of Escherichia coli formamidopyrimidine N-glycosylase: role of K155 in substrate binding and product release. Prog Nucleic Acid Res Mol Biol. 2001;68:223-34. PMID:11554299
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Kropachev KY, Zharkov DO, Grollman AP. Catalytic mechanism of Escherichia coli endonuclease VIII: roles of the intercalation loop and the zinc finger. Biochemistry. 2006 Oct 3;45(39):12039-49. PMID:17002303 doi:10.1021/bi060663e
- ↑ Burgess S, Jaruga P, Dodson ML, Dizdaroglu M, Lloyd RS. Determination of active site residues in Escherichia coli endonuclease VIII. J Biol Chem. 2002 Jan 25;277(4):2938-44. Epub 2001 Nov 15. PMID:11711552 doi:10.1074/jbc.M110499200
- ↑ Lavrukhin OV, Lloyd RS. Involvement of phylogenetically conserved acidic amino acid residues in catalysis by an oxidative DNA damage enzyme formamidopyrimidine glycosylase. Biochemistry. 2000 Dec 12;39(49):15266-71. PMID:11106507
- ↑ Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002 Feb 15;21(4):789-800. PMID:11847126 doi:10.1093/emboj/21.4.789
- ↑ Zaika EI, Perlow RA, Matz E, Broyde S, Gilboa R, Grollman AP, Zharkov DO. Substrate discrimination by formamidopyrimidine-DNA glycosylase: a mutational analysis. J Biol Chem. 2004 Feb 6;279(6):4849-61. Epub 2003 Nov 7. PMID:14607836 doi:10.1074/jbc.M310262200
- ↑ Golan G, Zharkov DO, Feinberg H, Fernandes AS, Zaika EI, Kycia JH, Grollman AP, Shoham G. Structure of the uncomplexed DNA repair enzyme endonuclease VIII indicates significant interdomain flexibility. Nucleic Acids Res. 2005 Sep 6;33(15):5006-16. Print 2005. PMID:16145054 doi:http://dx.doi.org/33/15/5006
- ↑ Shinmura K, Tao H, Goto M, Igarashi H, Taniguchi T, Maekawa M, Takezaki T, Sugimura H. Inactivating mutations of the human base excision repair gene NEIL1 in gastric cancer. Carcinogenesis. 2004 Dec;25(12):2311-7. Epub 2004 Aug 19. PMID:15319300 doi:10.1093/carcin/bgh267
- ↑ Tchou J, Michaels ML, Miller JH, Grollman AP. Function of the zinc finger in Escherichia coli Fpg protein. J Biol Chem. 1993 Dec 15;268(35):26738-44. PMID:8253809
- ↑ Tchou J, Michaels ML, Miller JH, Grollman AP. Function of the zinc finger in Escherichia coli Fpg protein. J Biol Chem. 1993 Dec 15;268(35):26738-44. PMID:8253809
- ↑ Kropachev KY, Zharkov DO, Grollman AP. Catalytic mechanism of Escherichia coli endonuclease VIII: roles of the intercalation loop and the zinc finger. Biochemistry. 2006 Oct 3;45(39):12039-49. PMID:17002303 doi:10.1021/bi060663e
- ↑ O'Connor TR, Graves RJ, de Murcia G, Castaing B, Laval J. Fpg protein of Escherichia coli is a zinc finger protein whose cysteine residues have a structural and/or functional role. J Biol Chem. 1993 Apr 25;268(12):9063-70. PMID:8473347









