Introduction
Isocitrate lyase is a lyase found in the proteome of multiple bacteria that oxidizes the hydroxl group of isocitrate and cleaves the substrate in two forming glyoxylate and succinate. Isocitrate lyase is a tetramer that is composed primarily of alpha helices and beta sheets with a unique structural phenomenon called "". This enzyme can be found within the cytosol of bacteria and is used in a variation of the citric acid cycle to help conserve energy by not using NADPH as an electron carrier and by reforming coenzyme-A earlier than in the normal citric acid cycle.
Isocitrate Lyase
Structure
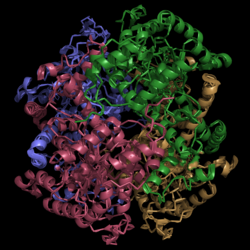
Figure 1. Crystal Structure of Isocitrate Lyase. Quaternary structure is comprised of four subunits forming an alpha/beta barrel.
Isocitrate lyase is a tetramer with 222 symmetry. Each subunit is composed of 14 alpha helices and 14 beta sheets which includes a total of 426 residues. These α helices and β sheets form an unusual α/β barrel seen in Figure 1. The α/β barrel contains a topology of (βα)2α(βα)5β, differing from the canonical (βα)8 pattern. Residues 184-200 and 235-254 connects the third and forth β-strands to their consecutive helices and form a that consists of a short five-stranded βsheet (β6,β7,β9,β10,β11) that lies on top of the α/β barrel. Isocitrate Lyase shows a resemblance to phosphoenolpyrvate mutase
Helix Swapping
A unique structural feature of this enzyme is a phenomenon called "".
Helix swapping is observed between two monomers to form stable dimers. The 12th and 13th helices of each monomer exchange three dimensional placement with the respective helices of the opposite monomer. Due to the 222 symmetry observed, only two dimers form than combine to form the observed tetramer. As a result of this structure, 18% of the surface of each monomer is buried within the protein.
Active Site Residues
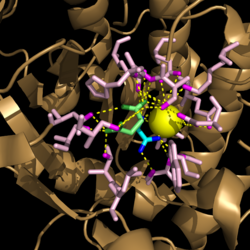
Figure 3. Active site residues hydrogen bound to a cofactor and the products of the catalyzed isocitrate reaction. Glyoxylate is shown in blue, succinate is shown in green, and the Mg
2+ cofactor is shown in yellow.
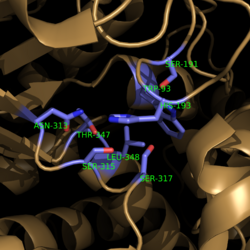
Figure 4. Active Site Residues. All eight active site residues necessary for catalysis of isocitrate are shown in slate. However, the protein shown is a C191S mutant of isocitrate lyase.
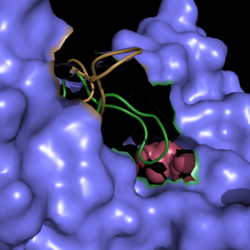
Figure 2. Active Site Loop Shift. Binding of the ligand to the enzyme results in a conformational shift that facilitates the breakdown of isocitrate. The active site loop unbound is shown in wheat and the active site loop bound is shown in green.
Regulation
Isocitrate lyase competes with isocitrate dehydrogenase, an enzyme found in the citric cycle, for isocitrate processing. The favoritism of one enzyme over the other is controlled by the phosphorylation of isocitrate dehydrogenase. This enzyme has a much higher affinity for isocitrate as compared to isocitrate lyase. Phosphorylation of isocitrate dehydrogenase inactivates the enzyme and leades to increased isocitrate lyase activity. [1]
Mechanism of Action
Image:TCA Cycle.png Figure 5. Citric Acid Cycle with Glyoxylate Shunt Pathway. In several bacterial species, there is a carbon conserving gloxylate shunt pathway that converts isocitrate to malate in two steps instead of the usual five steps.
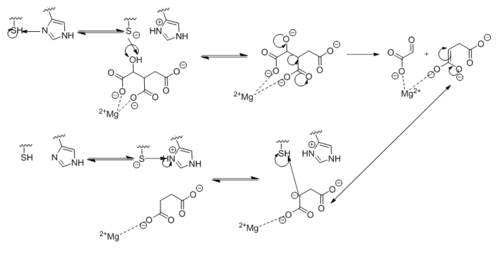
Figure 6. Observed Mechanism for the Breakdown of Isocitrate by Isocitrate Lyase. His193 shifts the pKa of Cys191 and removes its proton. This allows Cys191 to extract a proton from the hydroxyl group of isocitrate. The resulting oxyanion forms a carbonyl and forces the lysis of a C-C bond. Glyoxylate and the enol form of succinate are formed and stabilized with a Mg
2+ ion. The succinate enolate resonates and extracts the proton back from Cys191 to form succinate.
Disease Association
Clinical Implications
Mycobacterium tuberculosis is a respiratory infection that causes numerous fatalities throughout the world. It lives in organisms and feeds off of host cells, which indicate a variety of lipases exist within M. tuberculosis. Current drugs that are on the market now target a small number of bacterial processes like cell wall formation and chromosomal replication. Although several antibiotics exist, all of them target these same mechanisms of inhibition. These commonalities have led to the prevalence of different multi-drug resistant (MDR) tuberculosis strains. Due to the high level of resistance, finding a lasting treatment for MDR TB infections has become very problematic. Studies into new mechanisms of inhibition will be crucial to prevent widespread outbreaks.
Isocitrate lyase plays a key role in survival of M. tuberculosis by sustaining intracellular infections in inflammatory respiratory macrophages. Used in the citric acid cycle, isocitrate lyase is the first enzyme catalyzing the carbon conserving glyoxylate pathway. This glyoxylate pathway has not been observed in mammals and thus presents a unique drug target to solely attack TB infections.
Upregulation of the glyoxylate cycle has been seen for pathogens like M. tuberculosis that attack humans. [2] Furthermore, isocitrate lyase has been found to be essential for tuberculosis survival within hosts. [3]
Inhibitors
Due to the increased usefulness of this enzyme in propagating M. tuberculosis infections, specific inhibitors are being looked into as possible therapeutic targets for isocitrate lyase. Two such inhibitors that have already been identified are bromopyruvate and nitropropionate. Unfortunately, these molecules are non-specific and would also inhibit other enzymes essential for host function. [4] More research is needed to identify inhibitors that selectively target enzymes in the glyoxylate cycle.
Other 3D Structures of Isocitrate Lyase
- 1F61 Mycobacterium tuberculosis
- 1F8M Mycobacterium tuberculosis
- 1DQU Aspergillus nidulans
- 1IGW Escherichia coli
- 3IG3 Yersinia pestis
- 3I4E Burkholderia pseudomallei
- 3P0X, 3EOL, 3OQ8, 2E5B Brucella melitensis






{kind=link}