Overview
Mycobacterium tuberculosis NrdH is a small glutaredoxin-like protein involved in the electron transport chain in ribonucleotide reduction.[1]
Background
Mycobacterium tuberculosis Wikipediaresides in the lungs of a host and upon becoming active, results in symptoms such as chest pains, weakness, and intense coughing. Left untreated and unmanaged, TB can lead to death (1.5 million in 2013). The disease has a high co-morbidity with HIV/AIDS due to its immunocompromising tendencies. Tuberculosis is one of the most heavily studied diseases today. With over 9 million infections worldwide per year, the necessity for antimicrobial agents to combat emerging multi-drug resistant strands is imperative.
Structure
The structure of M. tuberculosis as determined by x-ray crystallography has 79 residues in a single polypeptide chain. . The active site (shown in green) is dominated by a disulphide bond between Cys-11 and Cys-14, which serves as the site of reduction by Theirodoxin reductase.
Many theirodoxin-like proteins have a similar active site region, denoted as the theirodoxin fold, which occurs directly before the disulfide bond. The residues in this region, denoted by letters CVQC, are the most highly conserved of all areas of the protein across multiple species. Exactly how this structure relates to function is somewhat debated. A Threonine-7 reside directly across the theirodoxin fold from the disulphide bond has been suggested to adopt two different conformations which differentially affect the redox abilities of the Protein. In the , the alcohol of the threonine side chain points towards the disulfide bond, engaging an ionic interaction between the two that prevents the Therodoxin Reductase from binding. Alternatively, in the , the alcohol points in the opposite direction, allowing sufficient space for the ligand to bind and reduction to occur.
The active site of the protein is stabilized through a network of hydrogen bonds involving the two highly conserved residues, CVQC and WSGFRP. The crystal structure shows that interactions with one water molecule is necessary for the proper coordination between the conserved motifs to occur. These hydrogen bonds orient the important residues in the most optimal position to promote oxidation and reduction.

Weblogo diagram showing highly conserved CVQC region of NrdH.
Another highly conserved residue is the WSGFRP sequence. This nonpolar sequence is found on the surface of the molecule and is exposed to solvent.
[2] [3] For this reason, it has been hypothesized that this sequence plays a role in the binding of thioredoxin reductase.
[4]
Arg-68 is responsible for the stabilization of the hydrophobic region of NrdH. Arg-68 has two distinct conformations. In the , Arg-68 is hydrogen bonded to His- 60 and Asp-59. When Arg-68 shifts to its
, it breaks it hydrogen bond with Asp-59. [5] This reduction in hydrogen bonding gives the hydrophobic region more flexibility and is thought to occur when NrdH is in its inactive state.
Function
MtNrdH has been identified as an electron carrier protein in ribonuleotide reduction. Ribonucleotide reduction uses an enzyme called ribonucleotide reductase (RNR) to make deoxyribonucleotides, which act as precursors to DNA synthesis. Three classes of RNRs have been identified; each class differs in cofactor requirement, structure, and oxygen dependence, but the general catalytic mechanism is conserved in all three classes.[6] Mycobacterium tuberculosis uses class I ribonucleotide reductase.
Class I RNR is further subdivided into class Ia and Ib. Both Ia and Ib reduce ribonucleotide 5’ diphosphate to deoxyribonucleotide 5’ diphosphate (NDP to dNDP). After ribonucleotide reductase performs the first round of reduction, RNR must be reduced again to reset the cycle. In class Ib, RNR is reduced by either glutadoxin or thiordoxin, which are first reduced by glutadoxin reductase and thiordoxin reductase, respectively.[7] In class Ib, RNR is reduced by NrdE, which is first reduced by NrdH. An important distinction between Ia and Ib is that Ia is present in eukaryotes, eubacteria, bacteriophages, and virus, but Ib is only present in eubacteria. [8]
[9]
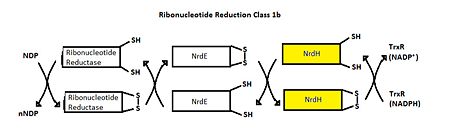
Ribonucleotide Reduction Class Ib general mechanism. The role of NrdH is highlighted.
Relevance
Like most NrdHs, MtNrdH is similar in sequence to glutaredoxins, but structurally similar to thioredoxins. MtNrdH also accepts electrons from thiordoxin reductase, a characteristic of thiordoxins, but not glutaredoxins.
[10] 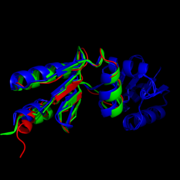
Structural comparison of NrdHs with "thioredoxin folds":
E. Coli NrdH (green),
C. ammoniagenes NrdH (blue),
M. tuberculosis NrdH (red)
Similar structures of NrdH have been isolated in other primitive species including E. coli, S. pyogenes, S. typhimurium, D. deserti, S. flexneri 2457T, and S. dysenteriae. In higher order multi-cellular organisms, however the NrdH protein is replaced by more complex glutaredoxins or theirodoxins. This observation leads some to speculate that NrdH is one of the very first ancestors in the ribonucleotide reduction pathway. If this is true, NrdH can be seen as a critical protein that allowed for the development of DNA-based life since deoxyribonucleotides could not have existed without the ribonucleotide reduction pathway. A better understanding of the evolutionary timeline of NrdH and similar proteins could shed greater light onto the RNA Wold Hypothesis, specifically describing the time frame of emergence of DNA based life.
Structural highlights
This is a sample scene created with SAT to by Group, and another to make of the protein. You can make your own scenes on SAT starting from scratch or loading and editing one of these sample scenes.Jmol reference [13] article describing Jmol [14]




{kind=link}