Introduction
The human cystathionine β-synthase (hCBS) is natively a homotetrameric enzyme (EC 4.2.1.22) catalyzing the following reaction (which is part of the biosynthesis of cysteine) using homocysteine and serine :
IMAGE
It is encoded by the CBS gene located on chromosome 21 (at position 22.3).
Structure
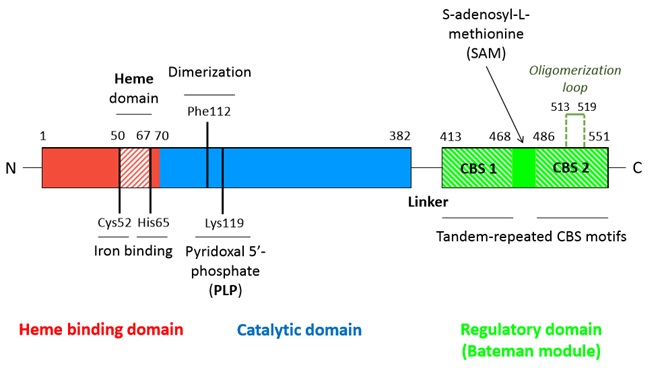
A CBS monomer is natively a 63 kDa protein made of 551 amino-acids and each of them binds two cofactors (the iron heme and the pyridoxal phosphate), as well as two substrates (homocysteine and serine). Hence, a CBS monomer contains (from N-terminal to C-terminal):
- a binding site located in a hydrophobic pocket (residues 50-67),
- a pyridoxal phosphate = (covalently linked to Lysine 119 amino group) situated in a (residues 70-382),
- a called Bateman module composed of two CBS domains (CBS1 and CBS2).
Cofactors
The heme is one of the two cofactors of hCBS.
It is bound in an hydrophobic pocket composed of the residues 50-67. The iron atom is hexacoordinated with the sulfhydryl group of Cys52 and the Nε2 atom of His65 (axial coordination) and with the four nitrogen atoms of the heme.
Although the heme is essential for activity in human CBS, its role still remains an enigma.
It is supposed to act as a redox sensor or as a way to facilitate a correct folding.
Quaternary structure
The hCBS is natively a homotetrameric enzyme. It is suggested that two monomers form a dimer, and then two dimers form a tetramer.
Two monomers shape a dimer through both hydrophobic and polar interactions within the catalytic core. Hydrophobic interactions particularly involve two Phe112 (one from each monomer) which interact with each other. There are no interactions between Bateman modules in a single dimer concerning hCBS.
It involves the Bateman modules as well as the catalytic cores of each dimer. Each (loop 513-519) of a monomer of one dimer interacts with the catalytic core of a monomer of the other dimer. Those loops interact within a crevace (shaped by α-helix 5-6-12-15-16 and β-strands 5-6) of the catalytic core. The tetramer is an inactive form of the enzyme.
This is the of the chain A of the protein.
This is the of the chain A of the protein.
This is the .
This is the of the protein.
This is the of the protein.
This is the .
Allosteric regulation
Autoinhibition (at the dimer scale)
The Bateman modules natively prevent the access of the substrates to the catalytic site (PLP cavity) through:
- hydrophobic interactions between residues I537, L540, A544 of the CBS1 domain of the of one monomer with residues I166, V189, V206, L210, I214 of the of the other monomer,
- hydrogen bounds between residues T460, N463, S466, Y484 of the CBS2 domain of the of one monomer with residues E201, N194, R196, D198 of the loop 191-202 of the of the other monomer.
These hydrophobic interactions combined with the hydrogen bounds network anchor the Bateman module of one monomer to the entrance of the catalytic core of the other monomer (close conformation), thus making it impossible for any substrate to access it.
Activation by S-adenosyl-L-methionine (SAM)
- SAM is the allosteric activator of the CBS. It binds in a region located between the CBS1 and CBS2 domains of the Bateman module which is solvent-exposed and has less hefty hydrophobic residues. Moreover, this region shapes a hydrophobic cage able to host the adenine ring. Moreover threonine (T535) and aspartate (D538) help stabilizing the ribose through hydrogen bounds and polar interactions.
- Binding of SAM to the Bateman module destabilizes the interactions which sustain the tetramer structure and thus triggers the dissociation of the tetrameric structure into two dimers.
- SAM fixation on the C-terminal regulatory domain entails the small rotation (or at least displacement) of the CBS1 and CBS2 of the Bateman module, thus distabilizing its interactions (hydrophobic interactions and hydrogen bounds) with the catalytic core of the other monomer. As a result, the Bateman module moves away from the catalytic core (this movement is all the more facilitated as the linker region is long enough and made of flexible residues). Loops 145-148, 171-174 and 191-202, previously involved in maintaining the close conformation through their interaction with the Bateman module, then relax and allow accessibility to the catalytic site (open conformation).
- SAM release leads to the return to the native inactive close conformation.
Disease
A tight control of CBS level and activity is crucial for optimal cognitive function :
- an overexpression of CBS (due to the localization of the CBS gene on the chromosome 21) is observed in the case of the Down Syndrome.
- on the contrary, a deficiency in CBS is linked to homocystinuria.
Relevance

