Background
Sex Lethal Protein (Sxl) is a splicing repressor in the male developmental pathway of sex determination of the common fruit fly, Drosophila melanogaster[1]. regulates alternative splicing pathways to promote the expression of female sex-linked proteins. In eukaryotes, splicing is carried out via the spliceosome, a ribozyme-protein complex which binds to the 5’ and 3’ splice sites. As Sxl is a splicing repressor, it prevents the binding of the U2AF and U1 subunits of the spliceosome at their respective splice sites, which represses the alternative splicing mechanism[2]. As a result, the fruit fly expressing Sxl will produce mRNA transcripts encoding proteins for the female developmental pathway[1].
Significance
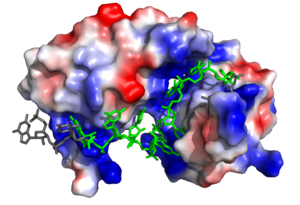
Figure 1. Three-dimensional representation of Sex-lethal protein showing the electrostatic surface and RNA ligand. Structure shown is
PDB:1b7f. Image created in PyMol.
The Sxl RNA splicing targets encode for the transformer (tra) and the male-sex lethal (msl-2) proteins. Tra is a splicing activator for the female developmental pathway, and Msl-2 modulates X chromosome application in male fruit flies. The mechanism for how Sxl targets these pathways differs slightly. In both mechanisms, Sxl occupies the 3' splice site and prevents U2AF from binding. This causes the U2AF splicing factor to bind at a downstream splice site encoding proteins in the female developmental pathway. In Msl-2 targeting, Sxl also blocks the binding of another regulatory splicing factor, TIA-1, and the U1 snRNP at the 5’ splice site. Sxl can also control its own splicing pattern to conserve female expression. It does so by binding to Exon 3 of its own RNA and creating an RNP complex to eliminate this exon. After removal of Exon 3, Sxl becomes active and female expression is maintained.
Structure
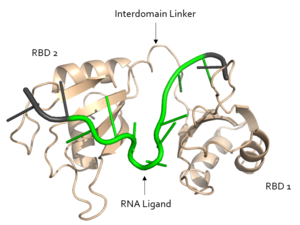
Figure 2. Structural overview of Sxl. RNA ligand colored in green is recognized and bound, while RNA ligand colored in grey is not bound. Image created in PyMol. Structure shown is
PDB:1b7f.
Sxl is composed of two asymmetric RNA binding domains (RBD1 and RBD2) which recognize a poly-uridine site in the pre-mRNA transcript[1]. Each RBD is comprised of two alpha helices and one antiparallel four-stranded β sheet[1]. The β sheets face each other, lining the electropositive V-shaped cleft[1]. The inter-domain linker forms a distorted 310 helix which helps form the V-shaped cleft into which the pre-mRNA sequence binds[1][3]. Sxl binds to UGUUUUUUU sequence of GUUGUUUUUUUU in tra. RBD1 binds U6-U11 and RBD2 binds U3, G4, and U5. Although the two RBDs do not interact with each other, this nine-ribonucleotide sequence must be recognized continuously to prevent U2AF from binding at the 3’ splice site[1]. The binding of Sxl to the pre-mRNA occurs in an electropositive pocket due to extensive interactions with the RNA phosphate backbone and negatively charged residues[1]. Since Sxl binds primarily with the phosphate backbone, the protein residues are not highly conserved.
Alternative Splicing Pathways
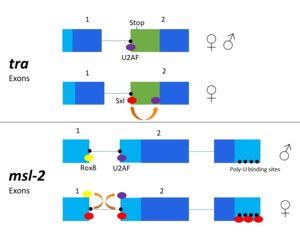
Figure 3. 2-dimensional representation of alternative splicing repression by Sxl on the
tra and
msl-2 genes.
The alternative splicing pathways of Sxl differ, but both involve repression at the 3' splice site. The tra expression pathway only involves the 3' splice site, while the msl-2 pathway involves both the 3' splice site and the 5' splice site. Both mechanisms cause U2AF binding downstream (Fig. 2)[3]. U2AF is a more general splicing factor than Sxl, preferring cytidine-containing poly-uridine pre-mRNA sequences, so Sxl binds to the guanosine-containing pre-mRNA with a 104-fold greater affinity.
Autoregulation
Sxl is capable of autoregulation of its expression[3]. The Sxl gene is transcribed in male flies, but the inclusion of exon 3 results in a premature stop codon, producing an inactive, truncated protein. The same Sxl promoter is active in female flies, but an additional (briefly active) Sxl promoter produces a transcript with exon 3 removed, producing an active Sxl protein which will initiate other female-specific splicing cascades[3].
Tra
In alternative splicing of the tra gene, Sxl binds at the 3' poly-uridine site. This causes U2AF to bind downstream and the spliceosome transcribes the following exon[2]. In the absence of Sxl, the normal gene for male development is transcribed. The exon contains a stop codon which results in a truncated, non-functional protein[3]. In the presence of Sxl, this exon is spliced, so the stop codon is skipped[3] (Fig. 3). This enables translation of an active tra protein[3].
Msl-2
The alternative splicing of msl-2 is reliant on Sxl binding to both the 5' and 3' splice sites. Sxl binds at the 3' splice site, replacing U2AF as in tra splicing. Sxl also competes with Rox8, which binds to the first intron. As a result, Sxl prevents splicing of the first intron of the msl-2 primary transcript. Sxl also binds to the poly- U sequences of the 3' UTR to repress translation (Fig.3)[4].
Structural Basis for Recognition of Poly-U Sequences
The ligand pre-mRNA sequence forms a characteristic loop at U5, U6, and U7. This interaction is stabilized by π stacking between the G4 and U5 nucleotides and residues , respectively. The nucleobases are exposed to residues on Sxl due to the 2’ endo conformation of all the nucleotides except for U8, which maintains a 3’ endo conformation.
The structural interactions with regards to the targeting of the 5' splice site and of its own mRNA transcript are much less understood than the competition of Sxl with U2AF at the 3' splice site.



