Hfq is a bacterial cytoplasmatic protein, first named for being a Host Factor for the Replication of the Qβ Phage RNA[1 - Schumacher 2002]. Hfq acts pleiotropically and impacts both the degradation and the translation efficiency of mRNAs [2 - Azam 2000], through the modification of sRNAs[3 - Wassarman 2001]. Besides, Hfq has a strongly conserved structure[4 - Kajitani 1994] and belongs to the LSm protein family, present in each of the three domains of life. Despite Hfq’s underscored importance, there is a wide spectrum of cellular functions, as growth rate and cell length, that are directly dependent of its chaperone function and have been discovered in the last century[5,6,7,8 - Artigos baixados], but many others remain unknown.
Structure
is a doughnut shaped homohexamer of roughly 8,9kDa, ~65Å diameter and ~23Å width, as described in S.aureus[1]. The action mechanisms of Hfq as a chaperone are based in its interaction with sRNAs. The best literature described functions among them are: the sequestering of ribosome binding sites (RBS); the inhibition of RBS-blocking mRNA regions, allowing translation; the stabilization of sRNAs which would otherwise be degraded; and the degradation of mRNAs. Hfq might also contribute to the degradation of certain mRNAs directly, but rarely. [9]
The which forms the homo-hexamer of Hfq has 77 residues[1] comprising one N-terminal α helix (α1), five beta sheets (β1-β5) and one variable region. An unstructured carboxy-terminal region, which may have >100 residues in certain species is also present[10 -Updegrove]. The fold of the beta sheets is characterized by two important , Sm1 and Sm2, used to clusterize the LSm protein family components considering their highlighted conservation throughout Life’s domains.
The most conserved amino acids of Sm and LSm proteins, including Hfq, are the aspartic acid 40 and the glycine 34, determining hydrophobic residues present in the Sm1 motif which maintain the highly distorted Sm1 fold. Tyr56 and Tyr63, highly conserved in the Sm2 motif, are fundamental for the interaction between subunits. The glutamine 8 and tyrosine 42 are highly conserved in Hfq proteins due to their role in uracil binding[1]. The highly variable C-terminus of Hfq proteins has no defined structure and its function is not clearly understood yet[10].
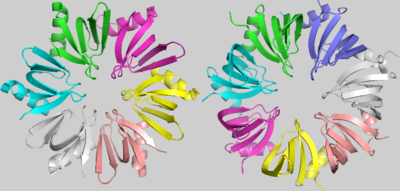
Hfq (Left) and SmAP1 (Right)
Although the strong conservation of some characteristic amino acids throughout the evolution of LSm proteins, there is an evident and important difference between the bacterial LSm proteins, including Hfq, and the archaeal and eukaryotic LSm protein members. Hfq, as denoted before, is an homo-hexamer, but archaeal and eukaryotic LSm proteins are formed by a homo-heptamer. images shows that this difference results from a shorter Hfq turn between its Sm1 and Sm2 motifs, named the “Variable region”. In addition, in archaeal and eukaryotic LSm proteins, the β3 and β4 strands are extended and the “Variable region” between them forms a little loop within its turn[1].
The is determined by hydrogen bonds between tyrosines present in the β4 and β5 sheets, the Tyr56 and the Tyr63, respectively. The interaction between residues from α1 and the turn between β2 and β3 also contributes to this conformation. The shorter “Variable region” present in Hfq, when compared to the heptameric LSm proteins, allows a closer rotation of the subunits, promoting the hexameric structure of Hfq[1].
The proximal face of Hfq interacts with U-rich 5′-AUUUUUG-3′ RNA sequences, forming a small around its pore. Residues such as Tyr42, Lys41 and Gln8 guarantee the bond with uracils and each subunit contacts one nucleotide, except the G residue that remains exposed out of the ring. However, in the predominantly , each subunit bonds with three nucleotides, forming a wider ring of RNA. In this case, A(A/G)N - N being any base - sequences are preferred.
Interestingly, the RNA binding promotes a conformational change in Hfq, highlighted by the pore diameter enlargement. The native protein shows a pore of roughly , but in case of active chaperone activity, the RNA bond promotes a pore diameter deformation to .
Structural highlights
This is a sample scene created with SAT to by Group, and another to make of the protein. You can make your own scenes on SAT starting from scratch or loading and editing one of these sample scenes.
You may include any references to papers as in: the use of JSmol in Proteopedia [1] or to the article describing Jmol [2] to the rescue. The Hfq protein is composed in its secondary structure.

