is a multidomain tumour suppressor protein involved in the regulation of various cellular processes, such as cell adhesion, migration or proliferation[1]. It is expressed in plethora of organs and tissues, e. g. cerebral cortex, bronchi or the gastrointestinal tract[2]. Germline truncation mutations of APC result in familial adenomatous polyposis, a hereditary form of colon cancer[3]. Additionally, loss of the C-terminal portion of APC is detected in about 80 % of sporadic colon cancers[4].
The overall structure of APC
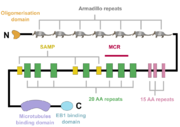
A schematic of the APC protein domain structure. MCR, mutation cluster region; SAMP, Axin-binding motif
The APC protein, its primary sequence encompassing 2843 aminoacids[5], consists of multiple domains, which enable it to interact with diverse partners. At the N-terminus, an oligomerisation domain is found, enabling the APC protein to oligomerise. It is followed by so called pre-ARM region and seven armadillo repeats, which form a groove for binding of a guanine nucleotide exchange factor Asef[6]. The central part of APC contains three 15 aminoacid long repeats followed by seven 20 aminoacid long repeats[1]. These motifs serve as binding sites for β-catenin[7]. In between the 20 aminoacid repeats, three SAMP regions are dispersed, enabling the interaction with Axin[1]. At the C-terminus, a basic domain responsible for binding to microtubules as well as EB1 interaction domain are present[8][1].
Interestingly, majority of somatic mutations occurs in so called mutation cluster region (MCR) between codons 1286 and 1513[9].
The physiological functions of APC and their implications for colorectal cancer onset and progression
Regulation of cell adhesion and migration
The seven armadillo repeats (ARM) together with the so-called pre-ARM region adjoining them at the N-terminus are essential for binding the guanine nucleotide exchange factor Asef[6]. In the absence of APC, Asef adopts an autoinhibited conformation, which prevents it from interaction with the small GTPase Cdc42[10]. Upon APC binding, the autoinhibited conformation of Asef is disrupted and the binding site for Cdc42 is made accessible[6]. Interaction with Asef leads to the exchange of GDP for GTP in the Cdc42 protein, which in turn modulates adherent junctions and contributes to enhanced cell motility[11][10][6]. In colorectal cancers, the truncated version of APC with preserved pre-ARM and ARM domains constitutively activates Asef and hence Cdc42[12]. This leads to extracellular matrix remodelling and promotion of adhesion-independent growth and cell migration[13].
Interestingly, APC takes part in strengthening the adherent junctions through the regulation of cellular distribution of E-cadherin and β-catenin. Full-length APC leads to increased levels of E-cadherin at the plasma membrane and decreases the pool of nuclear β-catenin in favour of the cytosolic one, enabling adherent junctions to be formed[14]. On the other hand, the truncated form of APC lacking the β-catenin interaction motifs is unable of such actions[11].
Regulation of cell proliferation through the Wnt pathway
APC controls cell proliferation as a negative regulator of the Wnt signalling pathway. Together with Axin, GSK3-β, CK1, PP2A and SCFβ-TRCP E3-ubiquitin ligase, APC forms so-called destruction complex, whose role is to promote the degradation of β-catenin[15][16][17]. APC binds Axin through the SAMP regions in APC central part, stabilises it and enables its oligomerisation[18]. Moreover, it has been proposed that APC supports the disociation of phosphorylated (=marked for degradation) β-catenin from the destruction complex[18] and that it is also essential for the movement of the destruction complex towards the plasma membrane[17]. However, degradation of β-catenin is not the only way employed by APC to antagonise the Wnt signalling. β-catenin binding motifs in the central part of APC compete with other interaction partners of β-catenin, such as the transcription factor TCF, thus preventing the expression of pro-proliferative factors (cyclin D, c-myc)[19][20][21][22][23]. Additionally, APC was observed to facilitate the export of β-catenin from the nucleus, hence delivering it out of reach of its target transcription factors[24][25]. Truncated APC defective in β-catenin binding can not effectively control the Wnt pathway signalling either by promoting β-catenin degradation or by preventing it from interacting with transcription factors, which might lead to overactivation of cyclin D or c-myc expression and hence excessive proliferation[19][20][25][18]. However, the first β-catenin binding repeat is often preserved in truncated APC mutants in colorectal carcinomas, which has brought forth the hypothesis that preserving some level of β-catenin downregulation is necessary to balance the potentionally harmful outcomes of constitutive β-catenin activation[26][27][28].
Regulation of cell division
APC interacts with the plus end of microtubules and stabilises them[29]. During mitosis, it is involved in the regulation of spindle formation and correct chromosome attachment control[30][31]. C-terminally truncated APC is unable to participate in these interactions, which leads to defects in the mitotic spindle formation as well as in the chromosome segregation. This enhances the risk of additional mutations due to the chromosome instability[32][30][33][31][34].
Gain of function APC mutants
Structural insights into APC interactions
Activation of Asef
This is a sample scene created with SAT to by Group, and another to make of the protein. You can make your own scenes on SAT starting from scratch or loading and editing one of these sample scenes.

