This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
Sandbox Reserved 1750
From Proteopedia
Displayed</scene></scene>==DNA RECOGNITION BY GAL4: STRUCTURE OF A PROTEIN/DNA COMPLEX==
| |||||||||||
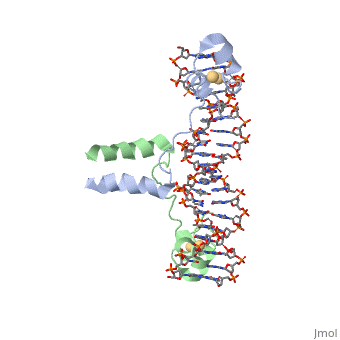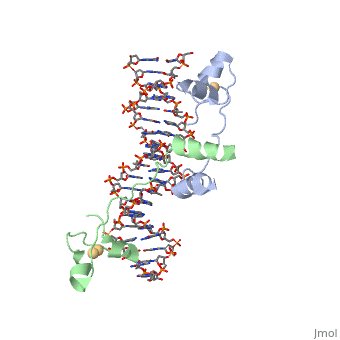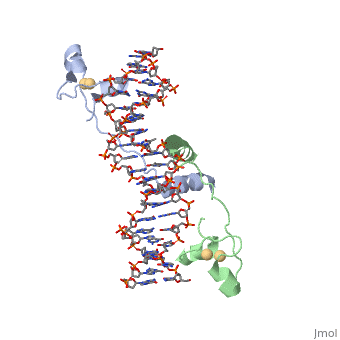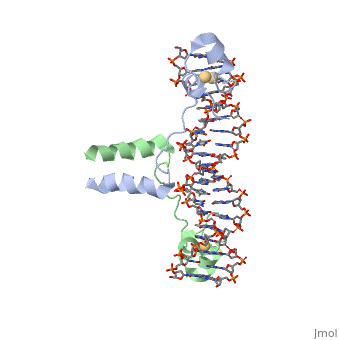

Contents |
STRUCTURE OF Cas9 IN STAPHYLOCOCCUS AUREUS IN COMPLEX WITH gRNA
Cas9 Overview
CRISPR is a bacterial immune response to bacteriophages to prevent subsequent infections and is a form of acquired immunity. Within the CRISPR system, Cas9 is a protein responsible for cutting the viral DNA rendering it inert. structure in Staphylococcus aureus (SaCas9) utilizes a single stranded guide RNA (sgRNA) to bind the target DNA that will be cut. Cas9 utalizes the sgRNA as an RNA guide for the target DNA sequence and binds complimentary to target DNA so Cas9 create a double stranded DNA break in the proper location. The target DNA must also have a PAM sequence to bind to Cas9 to be cut. The PAM sequence is a two factor authentication in junction with the sgRNA that tells the Cas9 to cut this portion of DNA. The main domains in the Cas9 are REC lob (residues 41–425) and NUC lobe (residues 1–40 and 435–1053). The NUC lobe contains RuvC, HNH, WEB, and PI domains (1). The REC lob is responsible for recognizing the nucleic acid, causing a conformational change in the HNH, and locking the HNH into the cleavage site (2). The RuvC nuclease (residues 1–40, 435–480 and 650–774) domain cleaves the strand of target DNA that does not bind the sgRNA (3). The HNH domain leaves the target strand of DNA bound to the sgRNA (3). The WEB domain (residues 788–909) is responsible for recognizing the sgRNA scafold and consists of twisted five-stranded beta sheet flanked by four alpha helices (4). The PI domain (residues 910–1053) recognizes the PAM sequence on the target DNA that is not complimentary to the sgRNA (5). There are also two linker domains, L1 (residues 481–519) and L2 (residues 629–649) that connect the RuvC and HNH (1). Furthermore, there is a phosphate lock loop (residues 775–787) that connect the WEB and RuvC domains. Cas9 has four main mechanisms that are important for successful cleavage including recognition of the sgRNA-target heteroduplex, recognition of the PAM sequence, recognition of the sgRNA scaffold, and endonuclease activity by HNH and RuvC.
Recognition of the sgRNA-target heteroduplex
The recognition of the sgRNA-target heteroduplex in Cas9 begins by inserting itself into the central channel between the REC and NUC lobes. The REC lobe interacts with the seed region of the sgRNA (C13-C20) as well as the PAM distal region (A3-U6) through the phosphate backbone. The seed region is in the A-form confirmation so it can bind the target DNA. The target DNA binds to the REC loop and RuvC domain for the proper conformation for base paring between the target DNA and sgRNA.
Recognition of the PAM seequence
For the recognition of the PAM sequence, the target DNA with the PAM sequence (5’-NNGRRN-3’) is bound to SaCas9 through bidentate hydrogen bonds as well as direct and water mediated hydrogen bonds through the major groove in the PI domain. The WED domain recognizes the minor groove phosphate backbone of the duplex.
Recognition of the sgRNA scaffold
The SaCas9 recognizes the sgRNA scaffold within the REC and WED domains. The WED domain contains five stranded beta sheets flanked with four alpha helices to allow binding of the repeat:anti-repeat duplex. REC lob binds the scaffold and secures it into the SaCas9.
Endonuclease Activity of Cas9
Finally, RuvC and HNH are in endonuclease activity. RuvC uses a two-metal ion mechanism of manganese to cleave the non-target DNA and causes a conformational change in L1. This conformational change leads to the phosphate group of the target strand to be cleaved by HNH. HNH includes a beta beta alpha metal fold and uses a one metal ion mechanism to cleave the target DNA.
Citations
1. Hiroshi Nishimasu, Le Cong, Winston X. Yan, F. Ann Ran, Bernd Zetsche, Yinqing Li, Arisa Kurabayashi, Ryuichiro Ishitani, Feng Zhang, Osamu Nureki, Crystal Structure of Staphylococcus aureus Cas9, Cell,Volume 162, Issue 5,2015,Pages 1113-1126,ISSN 0092-8674,https://doi.org/10.1016/j.cell.2015.08.007.
2. Palermo G, Chen JS, Ricci CG, Rivalta I, Jinek M, Batista VS, Doudna JA, McCammon JA. Key role of the REC lobe during CRISPR-Cas9 activation by 'sensing', 'regulating', and 'locking' the catalytic HNH domain. Q Rev Biophys. 2018;51:e91. doi: 10.1017/S0033583518000070. Epub 2018 Aug 3. PMID: 30555184; PMCID: PMC6292676.
3. Nishimasu H, Ran FA, Hsu PD, Konermann S, Shehata SI, Dohmae N, Ishitani R, Zhang F, Nureki O. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell. 2014 Feb 27;156(5):935-49. doi: 10.1016/j.cell.2014.02.001. Epub 2014 Feb 13. PMID: 24529477; PMCID: PMC4139937.
4. Morlot C, Pernot L, Le Gouellec A, Di Guilmi AM, Vernet T, Dideberg O, Dessen A. Crystal structure of a peptidoglycan synthesis regulatory factor (PBP3) from Streptococcus pneumoniae. J Biol Chem. 2005 Apr 22;280(16):15984-91. doi: 10.1074/jbc.M408446200. Epub 2004 Dec 13. PMID: 15596446.
5. Chen H, Choi J, Bailey S. Cut site selection by the two nuclease domains of the Cas9 RNA-guided endonuclease. J Biol Chem. 2014 May 9;289(19):13284-94. doi: 10.1074/jbc.M113.539726. Epub 2014 Mar 14. PMID: 24634220; PMCID: PMC4036338.
