The IgM BCR consists of six separate chains (Figure 1) that make up the three main
. A depiction of the IgM shows two heavy and two light chains that together form the
, or variable fragment at the top of the molecule where the antigen binding sites are located. The two heavy chains extend below the
which anchors the overall complex to the B cell. These regions are also labeled in Figure 1. The overall structure, expression, and function of the IgM BCR is strongly influenced by the
in which Ig α/β interactions as a heterodimer influence cell surface expression, receptor assembly, and effective signal transduction.
. In each domain, interactions between individual chains are important to understand the complex as a whole. All future 3D depictions showing these interactions ues the same PyMOL file
and will be as in Figure 1.
The IgM BCR is anchored to
B-cell membranes through the which is broken up into both extracellular and integral domains which sit on top of or span through the membrane, respectively (Figure 1). The extracellular region is primarily composed of
β-sheetswhile the integral region is composed of
α-helices. IgM BCR assembly requires dimerization of the
Igα and
Igβ subunits which embed within the B-cell membrane.
[4] The dimerizes within the extracellular region via a . Additional dimerization occurs within the integral region via a hydrogen bond; the residues involved have not been confirmed. Although the mechanism of disulfide bridge formation is still unknown, via
N-linked asparagine glycosyl groups (NAGs) in the extracellular region of both the
Igα and and
Igβ chains is hypothesized to facilitate this process. The NAG groups are believed to be essential for the recruitment of
Chaperone proteins to optimize the folding process.
[6] Past studies with human and viral proteins have shown that the presence of NAGs not only facilitate the rapid formation of disulfide bridges, but also ensure correct location.
[7],
[8] The recruited chaperone proteins will remain bound to the
Igα and
Igβ subunits until dimerization occurs.
[5]
After Igα and Igβ dimerization, the transmembrane helices of the heavy chains can embed within the B-cell membrane and intertwine with the Igα and Igβ chains. [4] The side chains of this , made up by the alpha, beta, and heavy chains (A/B), are primarily hydrophobic; this allows for interactions with the hydrophobic tails in the phospholipid bilayer. The four helices (Figure 2) are intertwined and primarily held together through interactions between the ; however, a a few polar residues are included which allow for additional interactions with the polar residues on the Igα and Igβ chains. [5]
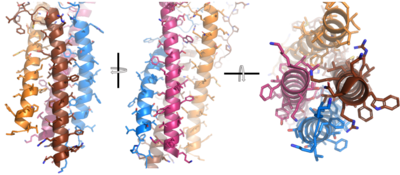
Figure 2. 4-pass integral helix. PyMOL image of the integral helices in IgM BCR (PDB:7xq8). The structure is shown rotated on the x and y axes to illustrate how the chains are intertwined. Side chains are shown as sticks. Brown=Ig alpha, orange=Ig beta, pink=heavy chain A, blue=heavy chain B.
Within the transmembrane region, heavy chain A and heavy chain B associate (Figure 1) asymmetrically to facilitate intracellular signaling cascades. The transmembrane heavy chain interface allows them to pack together via Van der Waals contacts, but there are also prominent between each chain. More specifically, the hydroxyl group from Ser584 on heavy chain A donates a hydrogen bond to Ser584 and to Ser588 on heavy chain B. This creates a bifurcated hydrogen bond, essentially forming a “fork” between the two chains to help stabilize them and maintain the transmission of the signal once the cell is activated. Because transmembrane Ig molecules cannot efficiently initiate the signal cascade, they must associate with the Igα and Igβ chains within the BCR. [2]
Furthermore, both the Igα and Igβ chains have cytoplasmic tails that extend into the B cell (Figure 1). Each of these tails contain an ITAM region to facilitate signal transduction (Figure 4). [3] The structures of the ITAM regions have not yet been determined.
Fc Region
The constant region of IgM is made up of the two . These heavy chains form a bridge connecting the Fab region or variable region to the transmembrane region (Figure 1). They also act as a wire that the variable region can send a signal through to the transmembrane region as a mechanical change.
help hold the heavy chains and Igα/Igβ chains together in the extracellular portion of the transmembrane region. Because a conformational change occurs throughout the entirety of the IgM-BCR complex, the Fc region must be able to tolerate the contortion of the structure as the antigen binds. In constant region two, which is located at the start of the Fc region, heavy chain A and heavy chain B make a to stabilize the IgM-BCR and drive downstream signaling.
To maximize the Fc region’s signal transduction efficiency and Van der Waals contacts, constant region two of heavy chain A makes an asymmetrical association with constant region three of heavy chain B to create a . More specifically, Arg243 and Arg251 residues from heavy chain A donate three hydrogen bonds to Leu433, Thr431, and Asp376 residues on heavy chain B. Furthermore, Leu313 of heavy chain A accepts a hydrogen bond from Thr429 on heavy chain B. By packing together this closely, it allows the IgM antibody to transmit extracellular stimuli, upon an antigen binding, to the intracellular region to facilitate its biological functions. [3]
Fab Region
The of the antibody is where antigen recognition occurs upon binding (Figure 1). On each arm is one heavy (A/B) and one light (A/B) chain, both containing domains identical to their respective counterparts. Repeats of β-sandwiches form the constant and variable domains within the Fab region as antigen recognition occurs at the variable domain while the constant domain connects it to the rest of the IgM complex. Because the Fab region of IgM is poorly resolved, a structural analysis of an HIV neutralizing antibody called VCR01 was performed to approximate where an antigen would bind to at the . [9]
The IgM-BCR contains areas referred to as complementary-determining regions(CDRs), which are where the antigen makes with the antibody on the Fab domain. Figure 2 depicts this as a surface representation given that the specific residues within the antigen-binding motif are unknown.
Due to the poor resolution of the Fab region, specific side chain interactions between the heavy (A/B) and light (A/B) chains have not been determined. Based on the general structure of an antibody it is estimated that each β-sandwich contains one disulfide bridge with additional hydrogen bonds. [2] The poor resolution does not allow them to be visualized within the structure, but upon understanding the significance of cellular signals being transmitted down to the Fc region, the presence of disulfide bonds would allow the protein to maintain its stability during conformational changes that occur as a result of signal modulation. The shows how the four heavy and light chain β-sandwiches fit together. The Fab region heavy chains attach to the Fc region heavy chains, before continuing down into the intracellular domain to interact with the Igα/Igβ subunits. The light chains (A/B) however are only connected to the heavy chains (A/B) within the Fab region, thus have no contact with the Igα/Igβ heterodimer.
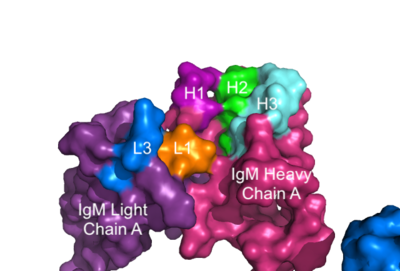
Figure 3. Surface Representation of IgM Antibody Binding Pocket. On one arm of the IgM antibody, the antigen makes contact with light chain A at the L1 and L3 complementary-determining regions. The antigen also makes contact with heavy chain A at the H1, H2, and H3 complementary-determining regions. The location of the complementary-determining regions were approximated using the structure of the VCR01 variable region and were visualized using Pymol.
Signal Transduction
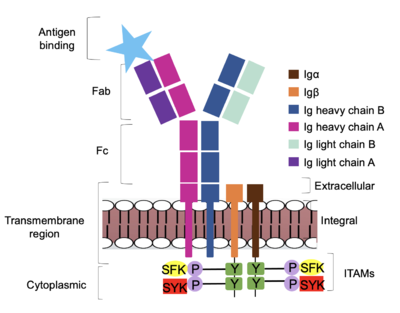
Figure 4. IgM Antibody Signal Transduction following Antigen Binding. The tyrosine residues on the intracellular end of Igα and Igβ helices are phosphorylated by potential tyrosine kinases: Splenic-tyrosine kinase and Src family kinase. Although the exact mechanism is unknown, the phosphorylation of Igα and Igβ activates the B cell and triggers intracellular downstream signaling.
Based on the structure of IgM, the diagram in Figure 4 depicts the initial process of B cell activation by the antigen binding to the antibody at the Fab region. The underlying mechanism for signal transduction is unknown due to the lack of specificity on which tyrosine residues in the
Igα and
Igβ domains are phosphorylated, but it is speculated to operate under what is known as the conserved assembly mechanism.
[3] Following an antigen binding, BCRs on the surface of the cell begin to cluster to cause the phosphorylation of the ITAMs located in
Igα and
Igβ. In its “off” state, the constant region 4 of
heavy chain B the extracellular components of
Igα and
Igβ. The antigen binding induces a conformational change to release the overlap and allow for clustering about the BCR. Now, in its “on” state, the phosphorylation of the
ITAM region within the intracellular tails of
Igα and
Igβ drives downstream kinase activity. As observed in Figure 4, the activated antibody can now continue to process of
intracellular signal cascading.
[3]




