The class 1 phaCs synthetize preferentially short chain PHAs, with 3 to 5 carbon monomers, and are composed by a , with molecular mass ranging from 63 to 73 kDA. (Chek et al., 2018; Neoh et al., 2022).
Thanks to the X-ray cristallography data from PhaCcn-CAT and PhaCcs-CAT, it was possible to categorize the class 1 phaCs in based on their molecular organization, made of two domains: The N-terminal domain and the catalytic C-terminal domain. The C-terminal domain carries the catalytic site, formed by the aminoacid triad (Cys-Asp-His) located deep within the hydrophobic cavity, that in the closed conformation is partially covered by the Cap subdomain.(Chek et al., 2018; Neoh et al., 2022).
N-terminal domain
The N-terminal domain has no defined function, and attempts to perform X-ray cristallography of this region have not been sucessful. Many studies have gathered evidence of possible roles that the N-terminal domain performs, such as: enzymatic activity efficiency, binding to PHA granules, substrate specificity, phaC expression, interaction with other PHA-related proteins and dimers formation and estabilization. Still, elucidation of its exact catalytic mechanism remains necessary.(Neoh et al., 2022).
C-terminal domain
Contrary to the flexible N-terminal domain, the C-terminal domain is relatively stable, making its crytalization process easier. Because of this, it was possible to obtain the C-terminal domain structure from PhaCcn-CAT and PhaCcs-CAT through X-ray cristallography, with resolution of 1.8 Å and 1.48 Å, respectively. The C-terminal domain has the catalytic site, with the aminoacid triad (Cys-Asp-His), the substrate entrance and the product egress tunnel.
The overall form of a phaC protein is that of a typical protein from the α/β-hydrolase-fold, with the C-terminal domain made of an α/β-hydrolase core subdomain and a Cap subdomain, corresponding to the Thr347-Pro471 residues in PhaCcn, and Thr319-Pro438 residues in PhaCcs. It is in the α/β-hydrolase subdomain that the entrance tunnel, the catalytic site and the product egress tunnel are located. This region seems to be preserved in phaCs.
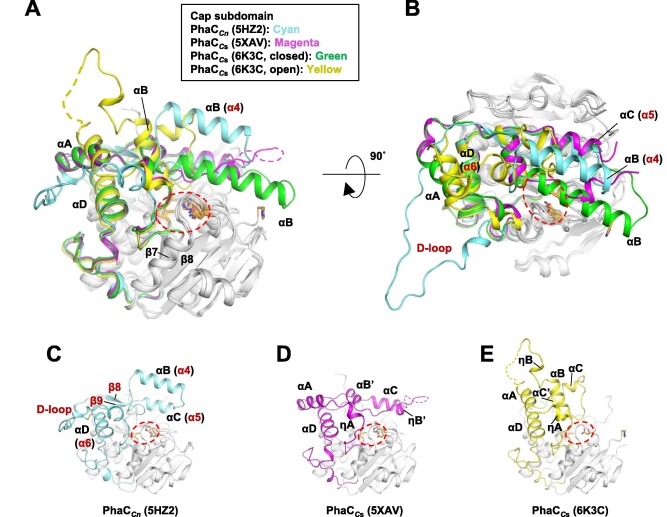
Regarding the Cap subdomain, the LID region is extremely dynamic and flexible, having an open or closed conformation based on structural changes. Because of this, the Cap subdomain, specially the LID region, is not as conserverd in the phaCs as the α/β-hydrolase subdomain. The Cap subdomain is located after the β7 sheet, and connects with the β8 sheet from the α/β-hydrolase core. In PhaCcn, the Cap subdomain is formed by three α-helixes (α4, α5 and α6) and two β-sheets (β8 and β9). Meanwhile, PhaCcs has six α-helixes (αA, αB, αC, αD, ηA and ηB').
The cristallography of PhaCcs-CAT bound to its substrate revealed the complex assymetric dimer structure of this enzyme. The phaC dimer form can be induced by the presence the substrate. Due to the dynamic and flexible properties, specially of the LID region, the Cap subdomain is paramount in the phaC dimer formation and regulation of substrate entry and product release, since it determines the protomer's movements, regulating the change between the closed form - homodimer- and the open form - heterodimer. (Neoh et al., 2022).
Secondary structure
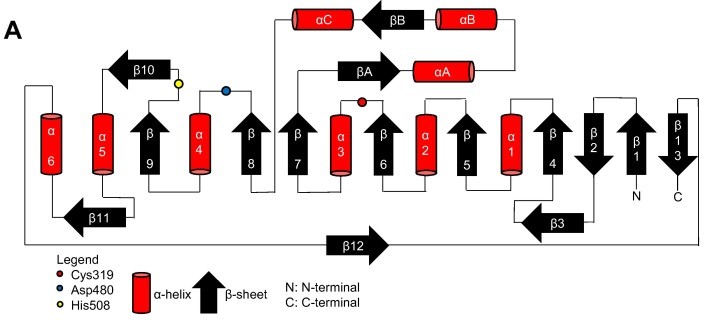
Topology diagram for the catalytic domain of the PhaCcn-CAT monomer. (Neoh et al., 2022).
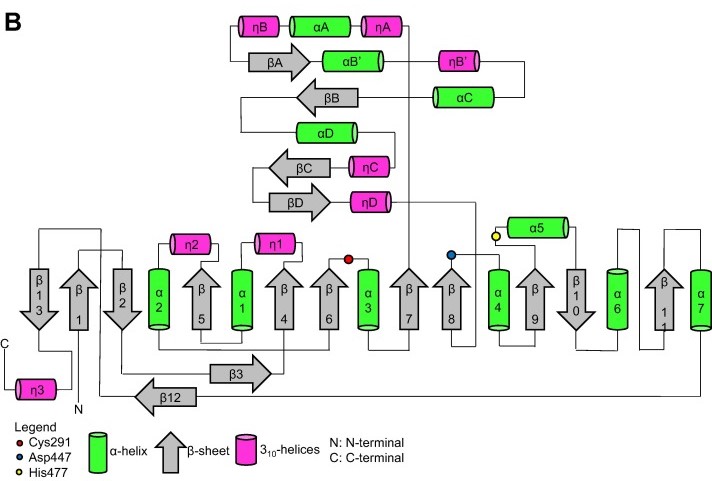
Topology diagram for the catalytic domain of the PhaCcs-CAT monomer. (Neoh et al., 2022).
Quaternary structure
Quaternary structure of PhaCcn-CAT and PhaCcs-CAT, with the Cap subdomain highlighted. (Neoh et al., 2022).