This old version of Proteopedia is provided for student assignments while the new version is undergoing repairs. Content and edits done in this old version of Proteopedia after March 1, 2026 will eventually be lost when it is retired in about June of 2026.
Apply for new accounts at the new Proteopedia. Your logins will work in both the old and new versions.
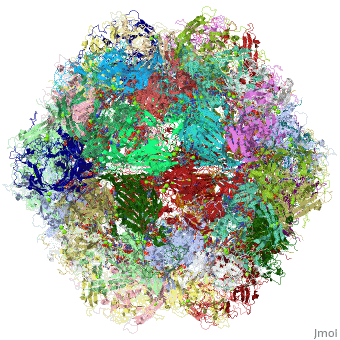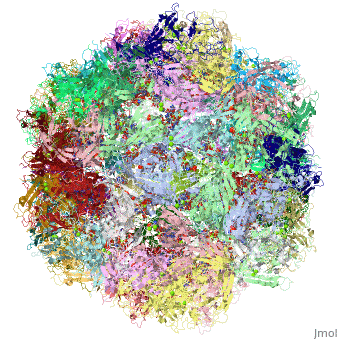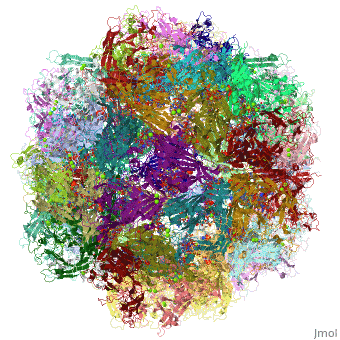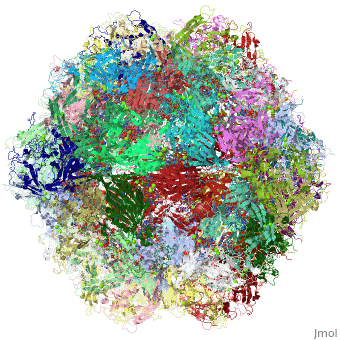
4dpv
From Proteopedia
PARVOVIRUS/DNA COMPLEX
Structural highlights
FunctionCAPSD_PAVCD Capsid protein self-assembles to form an icosahedral capsid with a T=1 symmetry, about 22 nm in diameter, and consisting of 60 copies of two size variants of the capsid proteins, VP1 and VP2, which differ by the presence of an N-terminal extension in the minor protein VP1. The capsid encapsulates the genomic ssDNA. Capsid proteins are responsible for the attachment to host cell receptor TFRC. This attachment induces virion internalization predominantly through clathrin-endocytosis. Binding to the host receptors also induces capsid rearrangements leading to surface exposure of VP1 N-terminus, specifically its phospholipase A2-like region and nuclear localization signal(s). VP1 N-terminus might serve as a lipolytic enzyme to breach the endosomal membrane during entry into host cell (By similarity). Intracytoplasmic transport involves microtubules and interaction between capsid proteins and host dynein. Exposure of nuclear localization signal probably allows nuclear import of capsids.[1] [2] [3] Evolutionary Conservation Check, as determined by ConSurfDB. You may read the explanation of the method and the full data available from ConSurf. Publication Abstract from PubMedThe DNA-containing capsid of canine parvovirus (CPV) is analyzed following atomic refinement at 2.9 A resolution. The capsid contains 60 copies of the capsid protein related by icosahedral symmetry. The atomic model has been extended from the first residue (Gly37) of the unrefined 3.25 A structure towards the N terminus. The electron density shows that approximately 87% of the capsid proteins have N termini on the inside of the capsid, but for approximately 13%, the polypeptide starts on the outside and runs through one of the pores surrounding each 5-fold axis, explaining apparently conflicting antigenic data. Analysis of potential hydrogen bonds reveals approximately 50% more secondary structure than previously apparent. Most of the additional secondary structure are in the 71 and 221 residue-long loop insertions between beta-strands E and F and G and H, forming subunit-bridging sheets that likely add specificity to assembly interactions. Structural analysis of the extensive subunit interactions around the 3-fold axes shows that assembly is a multistep process with loops intertwining following initial contact. Estimated free energies of association suggest that the formation of 3 and 5-fold contacts likely takes precedence over 2-fold interactions. Energies for initial association into trimers or pentamers would be similar, but the intertwining of loops about the 3-fold axis adds an additional large activation barrier to dissociation. Analysis of the surfaces of the assembled capsid shows a surprising lack of basic amino acids that might have been expected to interact with the negatively charged phosphoribose backbone of the DNA. Instead, uncharged polar and van der Waal's interactions predominate in the packaging of single-stranded DNA into the capsid. Canine parvovirus capsid structure, analyzed at 2.9 A resolution.,Xie Q, Chapman MS J Mol Biol. 1996 Dec 6;264(3):497-520. PMID:8969301[4] From MEDLINE®/PubMed®, a database of the U.S. National Library of Medicine. See AlsoReferences
| ||||||||||||||||||