Ashley Steere/Tobacco Etch Virus (TEV) Protease
From Proteopedia
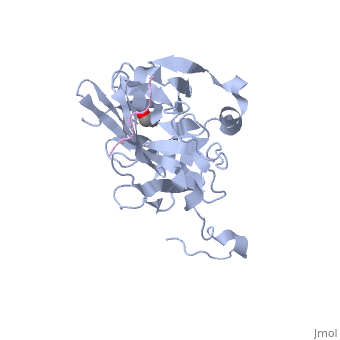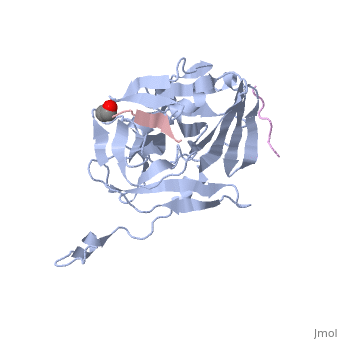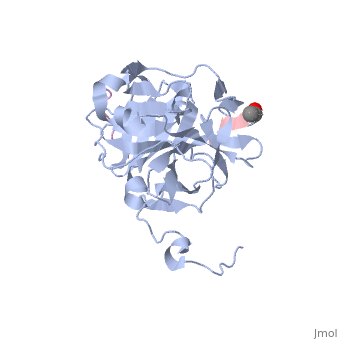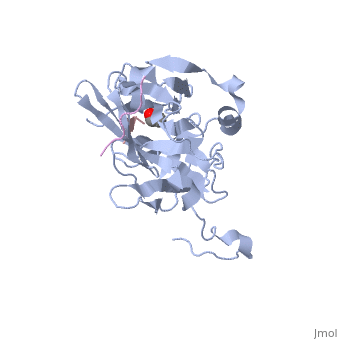
The tobacco etch virus (TEV) is a member of the Potyviridae family of positive-strand RNA viruses [1]. Unlike its name suggests, TEV can infect not only the tobacco plant, but a number of other plants including some flowers (baby's breath and zinnias) and fruits/vegetables (eggplant, peppers, and tomatoes). The 5' end of the RNA genome is covalently linked to a 49 kDa virus-encoded protein or VPg (viral protein genome-linked).The positive sense genome is translated by the host's ribosomes into a large polyprotein precursor that is cleaved soon after translation to form independent protein products [2]. The TEV nuclear inclusion a (NIa) protease (shown to the right co-crystallized with peptide product in purple, with the catalytic residues highlighted as green ball and stick models) [3] is a 27 kDa 3C-type cysteine protease, part of the VPg, and responsible for the processing of the original polyprotein into functional viral proteins. TEV protease resembles well-known serine proteases, such as trypsin and chymotrypsin, except that the TEV protease utilizes the nucleophilic thiol of the active site cysteine residue, as opposed to the serine hydroxyl used in serine proteases. Ultimately, the biological importance of the TEV protease requires that the enzyme have very stringent sequence specificity to ensure proper production of viral proteins, and it is based on this stringency that the TEV protease has increasingly been used to remove affinity tags from recombinant proteins. See also Tobacco Etch Virus (TEV) Protease.
Structure of TEV ProteaseTEV protease is classified as an all β protein which adopts a antiparallel β-barrel fold, typical of trypsin-like serine proteases, where the β sheet in the first domain folds to form an antiparallel (cyan) and the β sheet in the second domain is open (red). Like typical serine proteases, the β-barrel contains a motif. Located at the interface between the two domains is the , composed of His46, Asp81, and Cys151. A structural comparison with related proteins reveals that the TEV protease fold is most similar to that of other 3C-type cysteine proteases from hepatitis A virus and rhinovirus, which serve a similar function as the TEV protease in their respective viruses. However, although the overall fold of TEV protease and these related proteins is indeed very similar, the actual atomic coordinates are very different, with the root mean square deviation for α carbons between 2.4 to 3.5 Å [3]. Substrate SpecificityThe canonical recognition site of the TEV protease is the seven amino acid sequence ENLYFQ/G, with cleavage occurring after the glutamine residue. Although quite specific, TEV protease does have a tendency to undergo self-cleavage at a (KVFM/S) which follows residue 218, yielding a truncated enzyme with diminished catalytic activity [4]. Based on biochemical and structural data, it appears that the C-terminal region of the protein is relatively unstructured and flexible, allowing the scissile bond between Met218 and Ser219 to come dangerously close to the active site, where it is readily cleaved. Crystallization of the catalytically inactive TEV protease mutant (C151A) reveals that the of the protease (shown in yellow, active site residues in green) actually binds tightly within the active site in the absence of natural substrate. It is proposed that after cleavage at the non-canonical site, the free C-terminal peptide comprised of residues 219-242 of the TEV protease, binds within the active site of the enzyme in some type of auto-inhibitory mechanism [5]. This C-terminal peptide has no known counterpart in any other trypsin-like proteases and it is believed that in the presence of an appropriate substrate the C-terminus would be displaced from the active site, thus re-activating the enzyme. Mutation of Ser 219 to a number of other amino acids (Asp, Val, Pro) functions to limit the mobility of the peptide bond between residues 218 and 219, and has led to the production of TEV protease constructs resistant to this auto-proteolytic/auto-inhibitory activity with no affect on the normal catalytic function. LinksTo view available crystal structures of TEV protease please visit the following: References
| ||||||||||||||||||