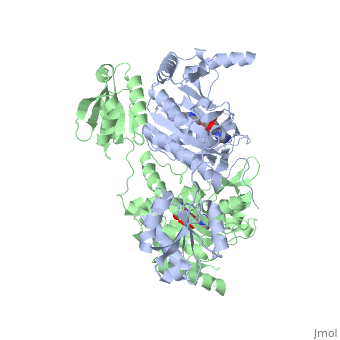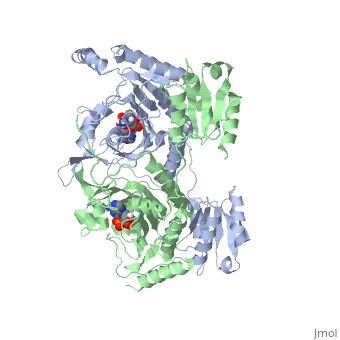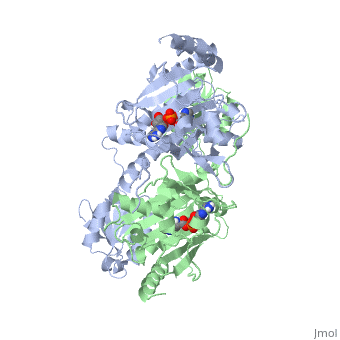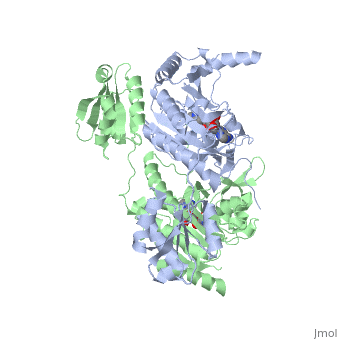
Histidyl tRNA Synthetase (HisRS) is a 94kD that belongs to the class II of aminoacyl-tRNA synthetases (aaRS). Aminoacyl-tRNA synthetases play a key role in protein synthesis and are classified as ligases. Aminoacyl tRNA synthetases use high energy ATP to attach a specific amino acid to its cognate tRNA[1]. This family of enzymes have been partitioned into two classes, containing 10 members, on the basis of sequence comparisons[1]. Class I and class II enzymes differ mainly with respect to the topology of the catalytic fold and site of esterification on cognate tRNA[1]. Class I aaRS enzymes contain a conserved Rossmann fold catalytic domain and often monomeric. Furthermore, class I enzymes attach the activated amino acid to the 2'OH of the tRNA molecule during catalysis, which then migrates to the 3'OH. Class II enzymes have a composed of anti-parallel (residues 1-325). Additionally, class II enzymes can be further divided into three subgroups: class IIa, distinguished by an N-terminal catalytic domain and C-terminal accessory domain (later shown to be the ); class IIb, whose anticodon binding domain is located in the N-terminal region; and class IIc, encompassing the tetrameric PheRS and GlyRS class II synthetases [2].
Histidyl-tRNA synthetases catalyze the transfer of histidine to a histidinyl transfer RNA molecule (tRNAHis). As HisRS is a class II aaRS enzyme it attaches the amino acid histidine to the 3’OH of the terminal ribose of tRNA[3]. Histidine is often an important amino acid in the active site of other enzymes and is unique as it can behave as either an acid or a base. The overall secondary structure of a HisRS monomer consist of 20% beta sheets and 37% helical character. It is structurally classified by CATH as an α-β layered sandwich. The CATH hierarchy of structural classification is based on: Class, Architecture, Topology, Homologous superfamily. Each of HisRS contains a N-terminal catalytic domain, C-terminal anticodon binding domain, a , II, and III, an insertion domain, a HisA and HisB loop, as well as an insertion domain. These structural elements are essential in assisting the two step mechanism carried out to aminoacylate tRNAHis with high fidelity.
Substrate Specificity
The first crystal structure solved for histidyl tRNA-synthetase was of E.coli. To date structures of E.coli HisRS complexed with ATP, AMP, histidine, competitive inhibitor (histidinol), histidyl-adenylate, and 5’-O-[(L-histidinylamino)sulfonyl] adenosine have been extensively explored. Many residues involved in substrate specificity and binding were identified. Not only have residues important for substrate binding been identified but also residues essential for catalysis.
Histidine Binding Pocket
The active site of HisRS contains a composed of highly conserved found in distinct sequences motifs. First, the LV/AAGGGLDYY loop (or ) forms one wall of the binding pocket. This HisA loop is highly conserved and extends over a part of the active site[3]. Second, the glycine-rich β-strand (sequence AGGRYDGL preceding ) comprises the histidine binding pocket floor and wall. Finally, conserved side chains that make direct contact with histidine are Glu83 and Gln127 (), which contact the α-amino and α-carbonyl functional groups, respectively, and Glu131 (motif II) and Tyr264, which make hydrogen bonds to the Nδ and Nε, respectively, of the imidazole ring[3].
ATP Cofactor Binding
Many interactions are required to prepare adenosine triphosphate (ATP) for attack by a bound histidine molecule and encourage the magnesium pyrophosphate moiety to act as a leaving group. Residues in the β strands and the loop portion of motif 2 are important in ATP contacts for HisRS[4]. Generally, residues involved in are among the most highly conserved in the HisRS family and for the most part shared by all members in class II. The π-stacking interaction between the adenine ring of ATP and provides specificity in the binding of ATP. The recognition of the N6 amino group of ATP involves the main chain carbonyl of Tyr122. The ATP ribose 2’ OH forms an additional contact with HisRS by hydrogen bonding with the main chain carbonyl of Thr281. There are four that mediate interactions with the triphosphate group of ATP. First, the conserved Arg113 forms a bridging interaction with the α and β phosphates of ATP in the crystal structure of e.coli HisRS complexed with histidinol [4]. Also, the γ phosphate forms salt bridges with conserved Arg121 and Arg311 in a complex with ATP. However, when HisRS is complexed with the adenylate, the Arg121 and Arg311 interactions are absent and adopt different conformations[5][4]. Furthermore, Glu115 also assists in the stabilization of the triphosphate group of ATP in a position such that it points back towards the adenine base. This conformation of ATP is evidently unique to class II aaRS[6].
The α phosphate of ATP interacts with conserved residue Arg113. The β and γ phosphates are neutralized by two coordinated magnesium ions that are positioned by water molecules and conserved Glu115[3]. The γ phosphate also forms interactions with conserved Arg121 and Arg311.
A Role for Coordinated Metal Ions
Thus far, residues of HisRS have been described for the binding of substrates histidine and ATP. However, HisRS also requires two magnesium ions to carry out catalysis. Most importantly, the β and γ phosphates of ATP are neutralized by two coordinated magnesium ions that are positioned by water molecules and conserved Glu115. Weak electron density, consistent with a bound Mg2+ ion, was observed in an electron density map for the HisRS:histidinol:ATP complex [4]. This particular Mg2+ ion coordinates the β and γ phosphates of ATP. Arnez et al. further defined the locations of magnesium ions by taking a crystal of HisRS:histidinol:ATP complex and soaking it in manganese(II) chloride MnCl2. These data showed that two Mn2+ ions coordinate the β and γ phosphates of ATP. Furthermore, interatomic distances between the Mn2+ principal ion and the β phosphate oxygen is approximately 0.5 Å, which would be expected to contribute to catalysis by weakening the bond between the α and β phosphates of ATP. In similar manganese soaking experiments with another classIIa aaRS, SerRS, the principal metal ion was shown to coordinate the α and β phosphates[7]. The functional role for the metal ion coordination between the α and β phosphates for SerRS is a metal-catalyzed mechanism for the adenylation reaction. Interestingly, Arg259 in the HisRS:ATP complex resides in the position occupied by the metal catalyst Mg2+, in classIIa SerRS. Arg259 and Arg113 serving in place of a Mg2+ ion is unique to HisRS compared to other classII aaRS. Other classII aaRS enzymes have conserved carboxylate groups to assist coordination of metal ions to carry out catalysis, while HisRS has in place residues Glu270 and Thr281 that have poor geometry for metal coordination but participate in the arginine salt bridge switch[4].(see figure below)
Mechanism of the Adenylation Reaction
below
Electrophilic Catalysis
The HisRS active site contains a highly conserved residue in the HisRS family, Arg259, that takes part in electrophilic catalysis for the adenylation reaction. This active site arginine residue is not present in other aaRS class II enzymes. As mentioned previously
are positioned to interact with the α-phosphate of ATP to assist in the adenylation reaction. Arg259 is positioned on the HisA loop to fix the α-carboxylate group of the histidine substrate as the [8]. Also, one ηN of the guanidinium group of Arg259 is positioned approximately 3Å from the α-phosphate of ATP while the other ηN hydrogen bonds with phenolic group of Tyr264. Thus, stabilization of the Tyr264 residue further enhances substrate binding allowing for the formation of a hydrogen bond to the Nδ of the histidine[3].
A comparison of in the HisRS:histidinol and the HisRS:adenylate complexes provides further structural information into how Arg259 may serve a role in catalysis. In the HisRS:histidinol complex a water-mediated interaction exists between Glu270 and εN of Arg259. However, in the HisRS:adenylate complex Glu270 moves to form a salt bridge with the guanidinium group excluding the water molecule. This movement serves as a salt bridge switch that may weaken the ionic interaction between Arg259 and the α-phosphate[4] and stabilize adenylate formation in the active site. Also, Arg113 as well as Arg259 are arranged to interact with of the histidyl-adenylate intermediate and stabilizes negative charge developed on the non-bridging oxygens α-phosphate during the transition state [3]. Evidence for Arg259 as critical residue in catalysis is further supported by mutational studies where a two or three log decrease in activity is observed when Arg259 is substituted with histidine [4] or other amino acids[9]. Utilizing Arg259 for catalysis is unique to HisRS as other class II aaRS enzymes, AspRS[10] and SerRS[7], use a divalent magnesium metal ion to coordinate the α-phosphate of ATP and serve as an electrophilic catalyst.
Mechanism of the Aminoacylation Reaction
below
Substrate Assisted Catalysis
The second reaction carried out by HisRS, , requires the decomposition of a mixed anhydride (the ) to form an aminoacyl ester on the 3’OH of tRNAHis. It was initially hypothesized that Glu83, acting as a general base, would improve the rate of this reaction. However, while Glu83 is in a favorable position in the active site to function as a base it is also situated to neutralize the α-amino group of the histidine substrate. Mutational analysis of Glu83[11] suggests that it does not act as a base but forms a salt bridge with the α-amino group of histidine, neutralizing the charge, and satisfying a critical electrostatic interaction.
The mechanism for transfer of the aminoacyl-adenylate intermediate onto its cognate tRNA is most easily explained by substrate assisted catalysis (SAC). During the substrate assisted catalysis the bond formation between the 3’OH of tRNAHis and the α-carboxylate carbon of the aminoacyl-adenylate occurs prior to the cleavage of the bond joining the α-carboxylate carbon to the axial oxygen of the α-phosphate. In the SAC mechanism the pro-S non-bridging oxygen acts as the base to abstract a proton from the 3'OH on the histidinyl tRNA. The functional role of the α-phosphate oxygen of the histidyl-adenylate was demonstrated by experiments utilizing phosphorothioate substituted histidyl-adenylate analogues. A sulfur atom in place of the either the Rp or Sp oxygen would be less basic and therefore reduce transfer of the histidine onto a tRNAHis molecule. The rates of aminoacyl transfer of the Rp thio-substitution adenylates were only 15-30 fold slower than the all oxygen adenylates[11]. However, the Sp thio-substitution reduced the rate of transfer at least 10,000 fold[11]. This thio effect supported the involvement of the non-bridging Sp oxygen in aminoacylation transfer chemistry. These results suggest that there is a stereospecific preference for the Sp-oxygen of the histidyl-adenylate intermediate in the HisRS active site.
Hence, in the SAC model the pro-S non-bridging oxygen acts as a base in abstracting the proton from the 3’ hydroxyl of A76 ribose. Then the A76 3’oxygen facilitates a nucleophilic attack on the carboxyl carbon of the histidyl-adenylate and breaks the mixed anhydride to release AMP. Also, Glu270 neutralizes Arg259 via a and elevates the pKa of the pro-S non-bridging oxygen. This concerted SAC mechanism rationalizes the considerable pKa difference between the 3’OH of tRNA (pKa =16) and the nonbridging Sp oxygen (pKa= -1)[11]. Therefore, as the bond between the tRNAHis nucleophile and the α-carboxylate is formed, the pKa for the 3’OH would be expected to drop sharply and the pKa of the nonbridging oxygen would tend to rise as the bond to the α-carboxylate lengthens and breaks[11].
Alternating Site Catalysis Model
The active site on one HisRS dimer has the potential to affect catalytic events in the second active site. Single turnover and stoichiometry studies of HisRS have indicated that one mole of adenylate is formed per mole of dimer. Furthermore, adenylate production is influenced by the presence of tRNA[12][11]. A unique model for explaining the coordination of catalysis between the two active sites of the HisRS dimer has been described by Guth and Francklyn in 2007 as Alternating Site Catalysis[12]. In this model histidine and ATP bind to one active site of the dimer to form the histidyl-adenylate intermediate. Following adenylate formation, tRNAHis will then bind to the this same subunit. A single round of aminoacyl transfer takes place in this active site, producing a “primed complex” with one aminoacylated tRNAHis per dimer. Instead of disassociating from the HisRS dimer it is theorized that the aminoacylated tRNAHis molecule remains bound while histidine and cofactor ATP are recruited into the second active site. The second active site will form the histidyl-adenylate and recruit another tRNAHis molecule. Finally, the aminoacylated tRNAHis from site one is released while the catalytic transfer of histidine onto tRNAHis occurs in the second site. Other class II enzymes that may also fit this asymmetric catalytic model include AspRS, SerRS, and ProRS. As an example, dramatically slowed kinetics for aminoacylation of tRNA is observed as a consequence of compromised accommodation for a second tRNA molecule in the E.coil AspRS:yeast tRNAAsp complex[13].
Histidinyl tRNA Recognition
below
The accuracy of protein synthesis is dependent upon the ability of aminoacyl-tRNA synthetases to specifically recognize the cognate tRNA and attach the appropriate amino acid. The identity nucleotides that define tRNA isoacceptor systems are primarily concentrated in the anticodon region and acceptor stems of tRNA molecules. These nucleotides provide functional groups that can be accessed by specific amino acid side chains of the aaRS enzymes[14],[15]. Also, tRNA identity and recognition can also emerge from the presence of modified bases such those commonly seen in the D-arm of tRNA molecules, dihydrouridine, or less common modification such as C5-methylation of uracil[16]. Key identity elements on E. coli histidine tRNA include the 5’ phosphate, G-1:C73 base pair in the acceptor stem and the GUG anticodon. Mutations of these identity elements diminishes aminoacylation in vitro[17] [18] [19]. Residues thought to be involved in the of these identity elements include: Arg123, Arg116, and Gln118. Substitution in Arg123 as a putative contact to the 5’phosphate, produced a 200 fold decrease in aminoacyl-transfer[12]. Similar kinetic defects in aminoacyl-transfer were also observed for Arg116 and Gln118 [12].
(see figure below)
Evolutionary Conservation
Structural Homology
During the course of evolution the catalytic domain has been the most preserved between prokaryotic and eukaryotic HisRS enzymes. Overall the sequence homology shared between prokaryotic, E.coli, HisRS and eukaryotic, S.cerevisiae, HisRS is only 28.5% in the catalytic domain, encompassing 235 residues [4]. The eukaryotic HisRS enzymes share much greater overall sequence homology, for example S.cerevisiae and H.sapiens HisRS have 43.5% shared sequence homology. Both eukaryotic and prokaryotic HisRS sequences share a conserved GExExxxG motif in the C-terminal domain. However, overall sequence homology shared between eukaryotic and prokaryotic C-terminal anticodon binding domain is minimal.
More recently a crystal structures for eukaryotic Trypanosomal brucei and Trypanosomal cruzi histidyl-tRNA synthetase was solved[20]. While both the human and trypanosomal HisRS sequences are part of the same eukaryotic branch on the HisRS phylogenetic tree[21] there is less than 30% sequence identity shared between them. Similarly, the sequence identity between trypanosoaml HisRS and bacterial HisRS is less than 30% [22]. Furthermore, many higher eukaryotes, more specifically mammals, have separate cytosolic and mitochondrial HisRS enzymes. The human HisRS genes, which arose from inverted gene duplication[23][24], HARS and HARS2 encode for the cytosolic and mitochondrial HisRS enzymes respectively. The gene products of HARS and HARS2 share approximately 79% sequence identity.