Crystal structures of forty- and seventy-one-substitution variants of hydroxynitrile lyase from rubber tree
Colin T. Pierce, Panhavuth Tan, Lauren R. Greenberg, Meghan E. Walsh, Ke Shi, Alana H. Nguyen, Elyssa L. Meixner, Sharad Sarak, Hideki Aihara, Robert L. Evans III, Romas J. Kazlauskas [1]
Molecular Tour
Beyond the Active Site: How Distant Residues Orchestrate Enzyme Function
Converting one enzyme into another might seem straightforward—just swap out the residues that directly contact the substrate, and voilà, you have a new catalyst. However, new structural studies reveal that this intuitive approach falls dramatically short. Researchers attempting to transform hydroxynitrile lyase from rubber trees into an esterase discovered that modifying only the 16 residues directly surrounding the substrate produced disappointing catalytic activity. The real breakthrough came when they expanded their engineering to include 40 or 71 mutations, creating variants that not only matched but exceeded the performance of their target esterase.
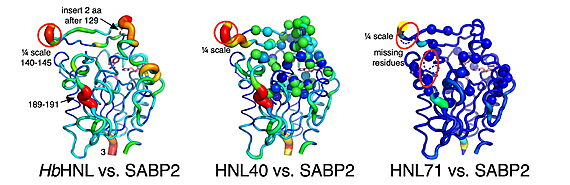
Displacement of Cα atoms (ΔCα) in HbHNL (PDB entry
1yb6), HNL40 (PDB entry
8sni), and HNL71 (PDB entry
9clr) relative to wt SABP2 (PDB entry
1y7i).
The crystal structures explain this dramatic difference in performance of this α/β-hydrolase fold enzyme[2][3][4][5] . The 40-mutation variant (HNL40) shows a partially restored oxyanion hole—a critical structural pocket that stabilizes the high-energy transition state during ester hydrolysis—while the 71-mutation variant (HNL71) achieves nearly perfect geometric matching with the target enzyme. Active-site & oxyanion hole of; [6], superposition of [7], superposition of , superposition of . Remarkably, both variants, i.e., HNL40 and HNL71, develop new tunnels connecting the active site to the protein surface, potentially providing escape routes for reaction products. These structural changes occurred with most mutations being located 6-14 Å away from the substrate-binding site.
These results reveal a sophisticated network where 'second-shell' residues, located beyond direct substrate contact, orchestrate catalytic efficiency through long-range effects. This finding suggests that creating truly efficient enzymes requires reengineering entire structural neighborhoods—a principle that could revolutionize approaches to enzyme design for biotechnology and medicine and help explain why natural enzymes maintain such complex architectures.
References
- ↑ doi: https://dx.doi.org/10.1107/S2059798325007065
- ↑ Ollis DL, Cheah E, Cygler M, Dijkstra B, Frolow F, Franken SM, Harel M, Remington SJ, Silman I, Schrag J, et al.. The alpha/beta hydrolase fold. Protein Eng. 1992 Apr;5(3):197-211. PMID:1409539
- ↑ Sussman JL, Harel M, Frolow F, Oefner C, Goldman A, Toker L, Silman I. Atomic structure of acetylcholinesterase from Torpedo californica: a prototypic acetylcholine-binding protein. Science. 1991 Aug 23;253(5022):872-9. PMID:1678899
- ↑ Chatonnet A, Perochon M, Velluet E, Marchot P. The ESTHER database on alpha/beta hydrolase fold proteins developments. Chem Biol Interact. 2023 Sep 25;383:110671. PMID:37582413 doi:10.1016/j.cbi.2023.110671
- ↑ The The ESTHER database website.
- ↑ Forouhar F, Yang Y, Kumar D, Chen Y, Fridman E, Park SW, Chiang Y, Acton TB, Montelione GT, Pichersky E, Klessig DF, Tong L. Structural and biochemical studies identify tobacco SABP2 as a methyl salicylate esterase and implicate it in plant innate immunity. Proc Natl Acad Sci U S A. 2005 Feb 1;102(5):1773-8. Epub 2005 Jan 24. PMID:15668381
- ↑ Wagner UG, Hasslacher M, Griengl H, Schwab H, Kratky C. Mechanism of cyanogenesis: the crystal structure of hydroxynitrile lyase from Hevea brasiliensis. Structure. 1996 Jul 15;4(7):811-22. PMID:8805565


