N-Butylisocyanide Oxidation at the [NiFe4S4OHx]-cluster of CO Dehydrogenase
Jae-Hun Jeoung and Holger Dobbek[1]
Molecular Tour
Ni, Fe-containing carbon monoxide dehydrogenases (CODHs) play an important role in anaerobic bacteria and archea by allowing them to grow with CO or CO2 as their sole carbon and/or energy source. The with ~ 130 kDa containing , called B, B’, C, C’ and D. Each subunit contains the and cubane-type [Fe4S4] . Another [Fe4S4] connects two subunits forming a covalent homodimer. The CODHs catalyze the reversible oxidation of CO to CO2 at the active site C-cluster, which is composed of [NiFe4S4OHx] (CO + H2O ↔ CO2 + 2e– + 2H+). In addition to the reversible oxidation of CO, CODHs are able to catalyze further reactions, such as the oxidation of H2 and the reductions of protons, 2,4,6-trinitrotoluene (TNT), and hydroxylamine, as well as the oxidation of n-butylisocyanide (n-BIC). N-BIC is a slow-turnover substrate of CODHs, whose oxidation occurs at the C-cluster.
The high resolution crystal structure of CODH-II from Carboxydothermus hydrogenoformans (dmin = 1.28 Å) revealed (the product of n-BIC oxidation), in which the (2yiv) rather than the (3b52) and states (3i39). , while . A superposition of the CODH-II structures with different ligands bound to the C-cluster reveals a flexible coordination and geometry for the Ni-Fe1 dyad, while the [Fe3S4] moiety of C-cluster remains unchanged in position.
, which was shown to be a binding place for Xe atoms in CODH/ACS (2z8y). Analysis of the CODH-II structure identified the presence of two different channels (see below), in which one is analogous to the channel identified by Xe-soaking in bifunctional CODH/ACS (conserved channel), while the other seems to be specific for monofunctional CODH (unique channel). The conserved channel in CODH-II is similar in position to that in bifunctional CODH, where the channel is extended to reach the A-cluster of ACS. Monofunctional CODHs have smaller side chains like Thr and Val, while Tyr617 and Phe634 block a passage of the unique channel to the protein surface in bifunctional CODH/ACS. Furthermore, the unique channel is completely blocked by residue His9 – Pro13 to prevent diffusion of gaseous substrate/product from the protein in bifunctional CODH/ACS. However, the channel of monofunctional CODH-II is directed towards the solvent, which is in line with its role to allow fast progress and egress of substrates and products from the active site to the outside of the enzyme, which has not been described previously.
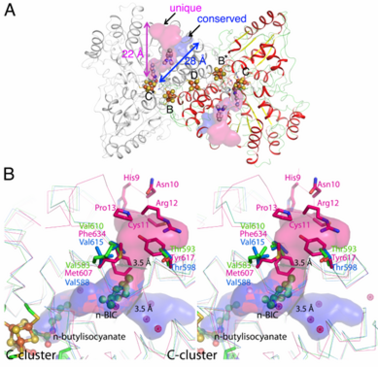
(A) Channels in monofunctional CODH-II
Ch. The channel into which the alkyl group of n-BIC is directed, shown in
pink color, is unique to monofunctional CODHs.
Blue space-filling model shows the channel found in both mono- and bi-functional CODHs. (B) Comparison of channels in mono- (CODH-II
Ch in
green from this study and CODH from
Rhodospirillum rubrum in
cyan (
1jqk)) and bi-functional (CODH from
Moorella thermoacetica in
pink from PDB-ID
2z8y) CODHs.
Pink-colored dots are Xenon atoms found in CODH from
Moorella thermoacetica.
PDB reference: NI,FE-CODH with n-butylisocyanate state, 2yiv.


