LiLac is a biosensor for lactate with a fluorescence-lifetime readout [1]. The fluorescent protein in LiLac is mTurquoise, a low pKa fluorescent protein commonly used in lifetime-contrast sensors. The ligand-binding protein in LiLac is the lactate-binding Cache domain of TlpC. Cache-domain-containing proteins sense extracellular nutrients to guide chemotaxis in bacteria; the N- and C- termini of the nutrient-sensing regions in Cache proteins (like the lactate-binding region of TlpC) are close in space, facilitating modular design. The combination of this modularity and the large diversity of ligands that Cache domains can sense have made this domain family an attractive starting point for building sensors. The mTurquoise is split in the middle of its seventh β-strand (β7) and then inserted the extracellular domain of TlpC at this position [2]. By engineering the connections between split mTurquoise and TlpC using a microfluidics-based screen, we developed LiLac, a lactate biosensor with a robust ~1.2 ns decrease (−35%) in fluorescence lifetime as lactate binds.
LiLac has the following parts - , an N terminal domain, the lactate binding , a and .
The current report describes the structural aspect of LiLac biosensor (as a part of the course BI3323-Aug2025), and this is entirely adapted from the work entitled " State-dependent motion of a genetically encoded fluorescent biosensor” by Rosena et al [3].
Mechanism of action
When lactate binds, the fluorescent protein rotates ~150° relative to the TlpC domain and translates by 30 Å as shown in the figure that depicts the twist upon lactate binding.
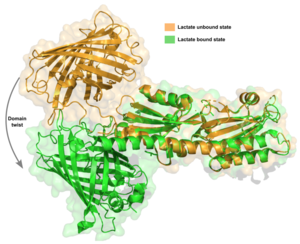
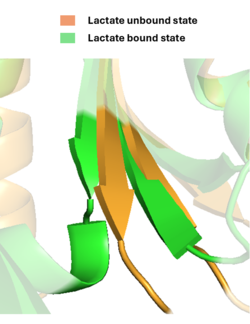
The first contributing feature is a “lid” that folds on top of lactate. The tip of this loop portion of LiLac in the absence of lactate, but it moves ~11 Å in the binding pocket.
Second, a at the back of the binding pocket, near its very C-terminus, shortens by one amino acid to accommodate lactate when it binds. This shortening is associated with a retraction of the TlpC “tail” in LiLac, which protrudes in the lactate-free form. Third, the tail undergoes a remarkable transformation in response to lactate binding, curling up into a short helical turn.
Mechanism for decreased lifetime upon lactate binding
The chromophore in our lactate-bound, low- lifetime structure lacked the seal normally seen in mTurquoise, almost certainly stabilizing the chromophore much less. In contrast, the chromophore in our lactate-free, high- lifetime structure was sealed shut. The “seal” for the mTurquoise portion of LiLac in a high-lifetime state was provided by the engineered , rather than the sequence that normally comprises the N-terminal half of β7; the N-terminal linker was largely disordered. The protein backbone of the C-terminal linker, as opposed to any of its specific amino acid side chains, is probably the “business end”.
In a high-lifetime state, F421 interdigitates tightly in between the outer face of the lactate-binding site and mTurquoise , in a small pocket consisting of the side chains of Q481, K483, L498, and F500.
In a low-lifetime state, F421 turns away from this interface, pointing instead between the stalk helix and the back of the lactate-binding pocket .
Summary
The engineered linkers between TlpC and mTurquoise are the key to the state-switching behavior in LiLac. At the level of the fluorescent protein, the structural studies of LiLac support a steric mechanism for how its fluorescence lifetime changes, where the chromophore is more “sealed” in a high-lifetime state than in a low-lifetime one. This change is described in light of a vibrational model for fluorescence-lifetime changes, but increased solvent access at this site could also lead to collisional quenching, additionally or alternatively explaining the lifetime change.


