Introduction
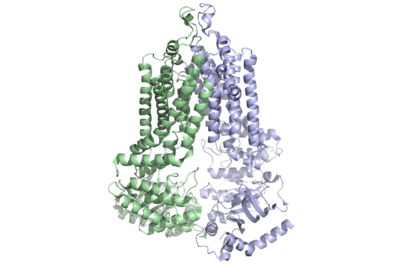
Figure 1: Multidrug-transporter ABCG2 is a dimer (6ffc).
The ABCG2 transporter protein is a notable transmembrane protein. It transports xenobiotic material out of cells in many tissues. ABCG2 belongs to the family of 48 transporter proteins called ATP-binding cassette transporters (ABC transporters). The ABC transporters differ from each other by their size structure, and ordering of domains. Ample evidence has shown a link between multi-drug resistance and the presence of ABC transporters in the plasma membrane of cells. This is important as multi-drug resistance is one of the major indicators of bad prognoses in cancer treatment. In fact, 19 of the 48 transporters of the ABC family have been shown to transport chemotherapeutic agents out of cells.[1] In recent studies, with cryogenic electronic microscopy (Cryo EM), the unique , inward facing nucleotide binding domain and of ABCG2 have been elucidated, among other features. These new discoveries have allowed for progress towards discovering the exact link between cancer and the ABC transporter family and have allowed for more effective drug treatment of cancer. This page will focus on the family of ABC transporters before delving into unique structural features of ABCG2 and finally describing the effects of this transporter on anti-cancer treatment.
ABC Transporter Family
In the 1990's, ABC binding cassette transporters became the subject of much discussion as many were found to have links to the inhibition of anti-cancer therapies. All 48 members of the family were studied and several structural aspects were found to be important to the characterization of transporters in this family. The first was the presence of two nucleotide binding domains (NBD) located in the cytoplasm of all cells which bound and hydrolyzed ATP, providing the necessary energy for transport of the substrate to occur. In all 7 subfamilies (A-G) of the ABC family, the NBD's are greatly conserved.[1] Each transporter of this family is made unique by the structure and form of their specific transmembrane binding domain (TMD). Each of the 48 transporters have 2 transmembrane domains which work to recognize and transport the substrates across the plasma membrane and out of the cell. The residues in the TMD provide the transporters each with specific substrates which they can transport. They also allow for the coupling of transport with ATP hydrolysis to transport molecules regardless of the concentration gradient.
Specific Members of the Family
As previously mentioned, 19 of the 48 members of the transporter family are involved in chemotherapeutic removal from the cell. Of these, three, , ABCG2 and were identified for further study and comparison of structure due to their function as multi-drug transporters.[1] The differences in their structures provided valuable information for scientific research into their substrate binding processes.
General Structure
The ABCG2 protein is comprised of a homodimer which each have two specific domains: one spanning the cell membrane and one involved with nucleotide binding.
Mechanism of Substrate Transport
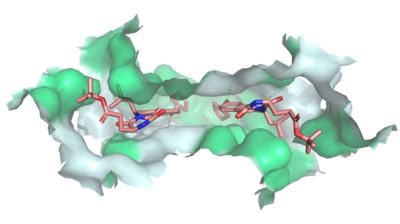
Figure 2: MZ29 bound to cavity 1 of ABCG2 (6ffc). Two MZ29 are shown in sticks and are colored by element. Hydrophobic interactions between the surface of cavity 1 and MZ29 are shown in green.
Multidrug Transporter ABCG2 is a that consists of two cavities separated by a . Cavity 1 is a binding pocket open to the cytoplasm and the inner leaflet of the plasma membrane. Its shape is suitable to bind flat, hydrophobic and polycyclic substrates.[2] Many of its amino acids residues form hydrophobic interactions with the bound substrate, as shown in green in Figure 1. Cavity 2 is located above the leucine plug. It is empty until a are bound to ABCG2.[3] Its (yellow is inter- and intra-molecular disulfides, golden is intra-molecular only) promote the release of the substrate from the cavity into the extracellular space.
One interesting feature of the NBD's is the fact that they remain in contact with one another even without a bound substrate. This makes the ABCG2 transporter unique and provides greater substrate specificity as the entrance to the transporter is not as globular as either ABCB1 or ABCC1. The entrance from the cytoplasm to the transporter is a hydrophobic membrane entrance lined by in both monomers.
Dimerization of ABCG2 was originally thought to be achieved with the help of the in each of the two domains but Cryo-EM showed that the motifs were on opposite sides of the protein.[2] [4]
Transmembrane Domain Stabilization
In the transmembrane domain is located the leucine plug that separates the first site of binding from the second site of binding for the substrate. Both domains are stabilized by several interactions. The first and most prominent feature of the TMD is EL-3. This loop which extends out from the transmembrane domain has been shown to be involved in stabilization of the TMD's. A member of the same subfamily, ABCG5/ABCG8, was shown to have that extended further into the extracellular space. This condensed helices is one of the defining features of ABCG2 transporter protein. The EL-3 of ABCG2 is stabilized by both intermolecular and intramolecular disulfide bonds at of each domain. ABCG8/ABCG5 do not have disulfide bonds present in EL-3 making it possible that intramolecular disulfide bonds in ABCG2 play a part in multiple drug transport as ABCG5/ABCG8 does not transport multiple chemotherapeutic drugs and has much less promiscuity of substrates than ABCG2. Even with the stabilization of the disulfide bonds, it has been discovered that without glycosylation at , ABCG2 does not mature and is ubiquitin tagged for degradation.
Function
ABCG2 transports a variety of , particularly flat, hydrophobic, and/or polycylcic molecules.[4] ABCG2 is found in multiple biological membranes, especially at blood barriers such as the brain barrier, testis barrier, and the blood-placental barrier. The prevalence of ABCG2 protects those tissues from cytotoxins. In addition to cytotoxin protection, ABCG2 secretes endogenous substrates in the adrenal gland, excretes toxins in the liver and kidneys, and regulates absorption of substrates.[5]
Fab-5D3
In the mid 2010's, the use of 2 antigen binding fragments allowed for high resolution images to finally be developed for ABCG2 transporter protein. With these recent discoveries, the understanding of this protein has greatly increased in the last several years. Before these images, ABCG2 had been unable to be recorded at high resolution as the domains were always moving. 5D3 Fab the two domains together preventing movement of the transporter from inward to outward facing. Fab binds at a 35 degree angle relative to the membrane plane which stops the 40 degree transition of the TMD from closed to open. Interestingly, complete arrest of ABCG2 in its inward facing state only requires one Fab bound but in experiments, two Fabs bound, each to both domains.
Polymorphic Changes
Several mutations have been shown to also decrease transporter activity. The first and most important of these is a point mutation of Glu 211 to Gln 211. This completely abolished activity of the transporter providing evidence of a significant role in the transport of substrates. Another important point mutation occurs at at , which becomes K141 and is the cause of gout. This mutation causes distortions of the protein's tertiary structure. The last mutation of note is at . Changes at this residue has been shown to change substrate specificity but as it is not in the binding pocket, this residue achieves this by allosteric interactions.
Disease
One of the causes for multidrug resistant cancers is the excretion of cancer drugs out of the cell, thereby decreasing the effective intracellular concentration. ABCG2, also known as the breast cancer resistance protein (BCRP), effluxes multiple chemotherapeutic agents such as mitoxantrone and camptothecin analogies, making the cancerous breast cells resistant to chemotherapy. Competitive inhibitors, such as , shut down ABCG2 to stop the efflux of cancer drugs in order to combat the resistivity of breast cancer. [4]
The ABC transporter family has been found as a prevalent piece of multi-drug resistant cancers and therefore became a popular target towards inhibition. Three generations of drugs were made in order to inhibit a similar protein from the same family, ABCC1 at its interior binding site including cyclosporine A (first generation), valspodar (second generation), and Elacridar (3rd generation). Importantly, cyclosporine A and Elacridar were found to inhibit both ABCC1 and ABCG2 and in one trial had success along with chemotherapy in the treatment of acute myeloid leukemia but because of either side effects or experimentation that was not able to be duplicated, this research was mostly shelved. The main issue in their failure to find a drug to inhibit this protein was the failure to develop a high-resolution structure of this protein with the technology available at the time of this drug development. Now, with high resolution images of the structures, fourth generation inhibitors are being developed with more success in inhibition without additional drug interactions or cell toxicity.
Relevance
As previously stated, by utilizing certain binders(), it is able to be stabilized for crystallographic imaging.[3] This has allowed researchers in the past decade to make advances based upon the greater understanding of its structure. Using these advances, inhibitors have been found to stop effluxion by ABCG2. Completely inhibiting this function, however, has residual effects on the excretory system. One such effect is decreased uric acid excretion in both the kidneys and the gut, which causes hyperuricemia . This results in an increased risk of uric acid crystal build-up, known as tophi formation, which causes a type of arthritis known as gout . Balancing the inhibition of ABCG2 will determine how to lessen these effects while continuing to combat cancer resistivity.[6]


